
Évaluation et Rapport de situation du COSEPAC sur le Renard gris Urocyon cinereoargenteus au Canada - 2015

- Table des matières
- COSEPAC - sommaire de l'évaluation - novembre 2015
- COSEPAC - résumé
- Résumé technique
- Préface
- Historique du COSEPAC
- Mandat du COSEPAC
- Composition du COSEPAC
- Définitions (2015)
- Description et importance de l'espèce sauvage
- Répartition
- Habitat
- Biologie
- Taille et tendances des populations
- Menaces et facteurs limitatifs
- Protection, statuts et classements
- Remerciements et experts contactés
- Sources d'information
- Sommaire biographique de la rédactrice du rapport
- Collections examinées
- Figure 1. Un renard gris (Urocyon cinereoargenteus) photographié en mai 2012, au cap Thunder, dans le nord-ouest de l'Ontario.
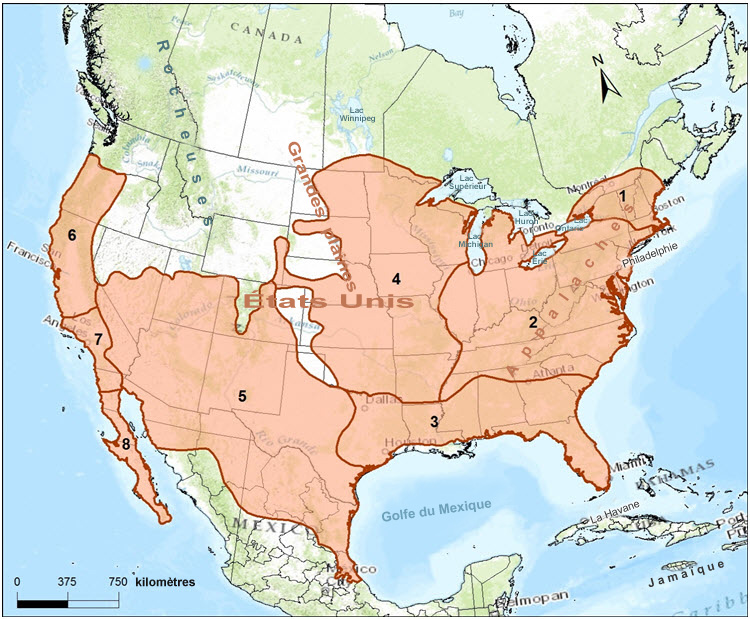
- Figure 2. Aire de répartition du renard gris (Urocyon cinereoargenteus) dans la partie continentale de l'Amérique du Nord, et localités de 8 des 15 sous-espèces. Le renard gris est présent du Canada à la Colombie, les sous-populations reproductrices canadiennes se trouvent dans les régions de Rainy River - Thunder Bay (nord-ouest de l'Ontario) et de l'île Pelée, en Ontario, et les sous-populations non reproductrices se trouvent au Manitoba et au Québec. Des individus qui se sont dispersés ont été signalés en Alberta et au Nouveau-Brunswick (non montrés sur la carte). Les sous-espèces qui sont montrées sont les suivantes : 1) borealis; 2) cinereoargenteus; 3) floridanus; 4) ocythous; 5) scotti; 6) catalinae; 7) californicus; et 8) peninsularis (Hall, 1981). Selon les analyses génétiques, les désignations des sous-espèces n'existeraient pas dans l'est de l'Amérique du Nord (Bozarth et al., 2011); dans le présent rapport de situation, une seule unité désignable est reconnue pour le renard gris au Canada.
- Figure 3 . Zone d'occurrence du renard gris (Urocyon cinereoargenteus), d'après les mentions récentes (c.-à-d. de 1993 à 2014) (indiquées par des étoiles) de l'espèce au Manitoba, en Ontario, au Québec et au Nouveau-Brunswick. Les mentions historiques (avant 1993) sont indiquées par des triangles.
- Tableau 1 . Dates des saisons de chasse et des saisons de piégeage et limites de prises de renards gris dans le nord-est des États-Unis et la région des Grands Lacs aux États-Unis.
- Tableau 2 . Cotes de NatureServe pour le renard gris aux États-Unis.
- Annexe 1 . précisions sur les observations de renards gris, par région, au Canada.
- Tableau A1. Mentions du renard gris au Manitoba. Source : Judge et Haviernick (2002), D. Berezanski (données inédites)
- Tableau A2. Mentions du renard gris dans le nord-ouest de l'Ontario. Source : Judge et Haviernick (2002), données du Centre d'information sur le patrimoine naturel (CIPN) et Van den Broeck (2014a). Les mentions non confirmées ne sont pas indiquées.
- Tableau A3. Mentions du renard gris à l'île Pelée (Ontario). Source : Judge et Haviernick (2002), données du Centre d'information sur le patrimoine naturel (CIPN) et Bowmanet al. (2013). Les mentions non confirmées ne sont pas indiquées.
- Tableau A4 Mentions du renard gris sur la rive nord du lac Érié, en Ontario. Source : Judge et Haviernick (2002), mis à jour à l'aide des données du Centre d'information sur le patrimoine naturel (CIPN). Les mentions non confirmées ne sont pas indiquées.
- Tableau A5. Observations du renard gris sur la rive nord-est du lac Ontario et du fleuve Saint-Laurent, en Ontario. Judge et Haviernick (2002), mis à jour à l'aide des données du Centre d'information sur le patrimoine naturel (CIPN).
- Tableau A6. Mentions du renard gris au Québec. Source : MFFP, données inédites.
- Tableau A7. Mentions du renard gris au Nouveau-Brunswick.
- Annexe 2. calculateur des menaces
- Calcul de l'impact global des menaces
- Tableau A8. Calculateur des menaces pour le renard gris.
- Menaces
COSEWIC
Committee on the Status
of Endangered Wildlife
in Canada

COSEPAC
Comité sur la situation
des espèces en péril
au Canada
Les rapports de situation du COSEPAC sont des documents de travail servant à déterminer le statut des espèces sauvages que l'on croit en péril. On peut citer le présent rapport de la façon suivante :
COSEPAC. 2015. Évaluation et Rapport de situation du COSEPAC sur le renard gris (Urocyon cinereoargenteus) au Canada. Comité sur la situation des espèces en péril au Canada. Ottawa. xiii + 61 p. (Registre public des espèces en péril).
Rapport(s) précédent(s) :
COSEPAC. 2002. Évaluation et Rapport de situation du COSEPAC sur le renard gris (Urocyon cinereoargenteus) au Canada - Mise à jour. Comité sur la situation des espèces en péril au Canada. Ottawa. vi + 34 p.
JUDGE, K.A., HAVIERNICK, M. 2002. Rapport de situation du COSEPAC sur le renard gris (Urocyon cinereoargenteus) au Canada - Mise à jour, in Évaluation et Rapport de situation du COSEPAC sur le renard gris (Urocyon cinereoargenteus) au Canada - Mise à jour. Comité sur la situation des espèces en péril au Canada. Ottawa. 1-34 p.
STEERS, S. 1979. COSEWIC status report on the grey fox Urocyon cinereoargenteus in Canada. Comité sur le statut des espèces menacées de disparition au Canada. Ottawa. 13 p.
Le COSEPAC remercie Jennie Pearce (Pearce & Associates Ecological Research) d'avoir rédigé le rapport de situation sur le renard gris (Urocyon cinereoargenteus) au Canada, aux termes d'un marché conclu avec Environnement Canada. La supervision et la révision du rapport ont été assurées par Graham Forbes, coprésident du sous-comité de spécialistes des mammifères terrestres du COSEPAC.
Secrétariat du COSEPAC
a/s Service canadien de la faune
Environnement Canada
Ottawa (Ontario)
K1A 0H3
Tél. : 819-938-4125
Téléc. : 819-938-3984
Courriel COSEPAC
Site Web COSEPAC
Also available in English under the title COSEWIC Assessment and Status Report on the Gray Fox Urocyon cinereoargenteus in Canada.
Illustration/photo de la couverture :
Renard gris - Photo : John Woodcock.
Renard gris
Urocyon cinereoargenteus
Le renard gris (Urocyon cinereoargenteus) présente un aspect typique d'un renard, mais son pelage est gris argenté, et une partie de la nuque, des flancs et des pattes est rougeâtre. Il possède une longue bande longitudinale foncée, sur toute la longueur du dos et de la queue. L'espèce est importante dans la phylogénie des Canidés parce qu'elle est considérée comme la plus basale de la famille des Canidés. Le renard gris est aussi l'espèce de Canidés la plus arboricole, et il peut partiellement faire tourner les os de la cheville pour faciliter la descente des arbres.
Le renard gris se rencontre généralement depuis le centre-sud du Canada jusqu'au nord de l'Amérique du Sud, et son aire de répartition prend de l'expansion vers le nord des États-Unis et l'est des États-Unis. Bien que l'espèce ait déjà été présente dans le sud de l'Ontario, elle ne se reproduirait actuellement que dans deux régions du Canada : la région de Rainy River -Thunder Bay (ci-après appelée le « nord-ouest de l'Ontario »), et la région de l'île Pelée, en Ontario. Dans le présent rapport, les individus observés dans le sud-est du Manitoba, au Québec et au Nouveau-Brunswick sont considérés comme se trouvant à l'extérieur des limites de l'aire de répartition de l'espèce ou comme des individus qui se sont dispersés ou des visiteurs ou, de manière plus appropriée, des individus dont la présence est occasionnelle et qui se sont dispersés à partir de populations voisines aux États-Unis et qui n'ont probablement pas établi de populations reproductrices dans ces régions du Canada.
La répartition de renard gris est étroitement liée aux forêts de feuillus. Le renard gris aménage sa tanière dans divers types de substrats, qui se trouvent habituellement dans des terrains couverts de broussailles denses et non loin d'une source d'eau. Il est considéré comme une espèce généraliste en matière d'habitat et tolère partiellement les perturbations humaines, bien qu'il soit plus discret que le renard roux (Vulpes vulpes) et qu'on le voit donc moins souvent que ce dernier.
De tous les Canidés d'Amérique du Nord, c'est le renard gris qui est le plus omnivore; il consomme des matières végétales, comme des fruits, à longueur d'année. Au Canada, le renard gris se reproduirait de la mi-février à la mi-mars, comme il le fait dans le nord-est des États-Unis. La plupart des individus femelles se reproduisent au cours de leur première année, et la portée compte 3 ou 4 petits par année. Le renard gris est généralement nocturne ou crépusculaire. L'unité sociale de base comprend un mâle adulte, une femelle adulte et leur progéniture, et ce groupe maintient un domaine vital. Le rapport des sexes des adultes est généralement 1:1, et il est présumé que le renard gris est monogame.
La taille des populations n'est pas connue. Bien qu'aucune étude quantitative à l'échelle des populations de renards gris au Canada n'ait été publiée, selon les rares mentions de l'espèce, il semble que le nombre d'individus matures présents au Canada serait inférieur à 110. Les populations de renards gris ont augmenté dans le nord des États-Unis, et l'aire de répartition a connu une expansion simultanée vers le nord, ce qui a entraîné des mentions supplémentaires de l'espèce au Canada.
La mortalité due au piégeage empêcherait l'établissement de sous-populations reproductrices dans une grande partie de l'aire de répartition canadienne du renard gris, à l'extérieur de l'île Pelée. Le facteur limitatif le plus important pour les populations de renards gris aux États-Unis est le piégeage. Bien que l'effet de la récolte au Canada n'ait pas été déterminé, la plupart des mentions dans la majeure partie des sous-populations sont liées au piégeage accidentel. En raison de la petite taille des populations, toute cause de mortalité importante, comme le taux de prédation élevé par le coyote et les maladies (y compris la maladie de Carré et la rage), pourrait devenir un facteur limitatif important. Dans les deux sous-populations pour lesquelles il existe des indices de reproduction, et principalement dans la sous-population du nord-ouest de l'Ontario, les menaces probables pesant sur le renard gris sont la mortalité due au piégeage et la mortalité routière. L'impact global des menaces est élevé.
L'espèce est classée par NatureServe comme très commune (G5) à l'échelle mondiale; cependant, au Canada, elle est classée dans la catégorie des espèces gravement en péril (N1). Le renard gris figure actuellement parmi les espèces menacées à l'annexe 1 de la Loi sur les espèces en péril (LEP). En Ontario, le renard gris a été désigné espèce menacée en vertu de la Loi de 2007 sur les espèces en voie de disparition de l'Ontario. Au Québec, il est considéré comme une espèce résidente occasionnelle et n'est ni classé ni inscrit. Au Manitoba, au Québec et au Nouveau-Brunswick, les renards gris observés sont considérés comme des individus qui se sont dispersés à partir des États-Unis et dont la présence est occasionnelle. Le renard gris serait présent, quoique rare, dans le parc national du Canada des Îles-du-Saint-Laurent; il a été observé dans la Réserve naturelle provinciale de la Pointe Fish, à l'île Pelée, et sa présence est mentionnée à l'occasion dans la région du parc provincial Whiteshell, dans le sud-est du Manitoba.
| Sujet | Information |
|---|---|
| Durée d'une génération (généralement, âge moyen des parents dans la population; indiquer si une méthode d'estimation de la durée d'une génération autre que celle qui est présentée dans les lignes directrices de l'UICN [2008] est utilisée) Selon la méthode 1 de l'UICN (âge moyen des parents), et un taux de survie annuel des juvéniles et des adultes de 0,31 et de 0,43 respectivement, un taux de fécondité de 3,7 et un âge des individus à la première reproduction de moins de 1 an. Les taux de survie estimés sont fondés sur les populations des États-Unis faisant l'objet de piégeage. |
Probablement 2 ans |
| Y a-t-il un déclin continu [inféré] du nombre total d'individus matures? Il n'existe aucune donnée sur les individus matures. Cependant, dans le nord-ouest de l'Ontario, il est possible que les observations augmentent, et la présence de femelles matures, d'une femelle en lactation et de couples indique qu'il y a probablement reproduction; cette population pourrait être en augmentation. La population de l'île Pelée pourrait être stable. Les mentions d'individus dans le sud-est du Québec augmentent, mais la plupart de ces mentions ne concernent probablement pas des individus matures. |
Peu probable |
| Pourcentage estimé de déclin continu du nombre total d'individus matures sur [cinq ans ou deux générations] (Voir ci-dessus.) |
Inconnu |
| Pourcentage [présumé] [d'augmentation] du nombre total d'individus matures au cours des [dix dernières années ou trois dernières générations]. (Voir ci-dessus.) |
Inconnu |
| Pourcentage [présumé] [d'augmentation] du nombre total d'individus matures au cours des [dix prochaines années ou trois prochaines générations]. Il est difficile de faire une projection parce qu'on ne connaît pas la gravité des menaces qui se produisent et que les données sur les individus matures sont limitées. La sous-population de renards gris dans le nord-ouest de l'Ontario devrait augmenter alors que la sous-population de l'île Pelée est probablement stable. Le nombre d'individus qui se déplacent jusqu'au Manitoba, jusque dans la partie continentale de l'Ontario, jusqu'au Québec et jusqu'au Nouveau-Brunswick augmentera probablement, et ces individus pourraient se reproduire dans l'avenir. |
Inconnu |
| Pourcentage [présumé] [d'augmentation] du nombre total d'individus matures au cours de toute période de [dix ans ou trois générations] commençant dans le passé et se terminant dans le futur. (Voir ci-dessus.) |
Inconnu |
| Est-ce que les causes du déclin sont clairement réversibles et comprises et ont effectivement cessé? Sans objet; les sous-populations augmentent, et il n'y a aucun signe de déclin de la reproduction (sauf dans l'est de l'Ontario où certaines mentions d'individus reproducteurs ont été effectuées dans les années 1950). |
S.O. |
| Y a-t-il des fluctuations extrêmes du nombre d'individus matures? Il existe peu de données sur les individus matures. |
Inconnu |
| Sujet | Information |
|---|---|
| Superficie estimée de la zone d'occurrence D'après la méthode du plus petit polygone convexe qui inclut seulement les superficies connues ou présumées renfermant des individus reproducteurs à l'île Pelée et dans le nord-ouest de l'Ontario. La valeur plus élevée inclut des superficies avec mentions récentes (c.-à-d. au cours des 20 dernières années) d'individus qui se sont déplacés au Canada, mais qui ne se reproduiraient pas. |
Zone d'occurrence : 35 173 km2 (625 517 km2) |
| Indice de zone d'occupation (IZO) [Fournissez toujours une valeur établie à partir d'une grille à carrés de 2 km de côté]. D'après les sites de reproduction possible à l'île Pelée et dans le nord-ouest de l'Ontario. La valeur plus élevée inclut des superficies avec mentions récentes (c.-à-d. au cours des 20 dernières années) d'individus qui se sont déplacés au Canada, mais qui ne se reproduiraient pas. |
IZO : 96 km2 (200 km2) |
| La population totale est-elle gravement fragmentée? Les deux sous-populations reproductrices sont isolées l'une de l'autre, mais la sous-population du nord-ouest de l'Ontario est connectée aux populations des États-Unis les plus proches en raison de la dispersion d'individus. Le degré d'isolement de la population de l'île Pelée des autres sous-populations n'est pas connu. |
Non |
| Nombre de localités (Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN (février 2014) pour obtenir des précisions sur ce terme.) L'île Pelée constituerait une localité parce que des individus existent dans une superficie relativement petite dans laquelle la menace liée au piégeage accidentel varie peu, alors que la sous-population du nord-ouest de l'Ontario subit une pression de piégeage qui varierait dans l'ensemble de la région. |
Inconnu; probablement > 10 |
| Y a-t-il un déclin continu [prévu] de la zone d'occurrence? La zone d'occurrence a augmenté avec l'ajout de mentions provenant du nord-ouest de l'Ontario. |
Non |
| Y a-t-il un déclin continu [inféré] de l'indice de zone d'occupation? L'IZO a augmenté depuis 2002 en raison de l'établissement d'une sous-population reproductrice dans le nord-ouest de l'Ontario. |
Non |
| Y a-t-il un déclin continu [inféré] du nombre de populations? Le nombre de sous-populations a augmenté depuis 2002 en raison de l'établissement d'une sous-population reproductrice dans le nord-ouest de l'Ontario. |
Non |
| Y a-t-il un déclin continu [inféré] du nombre de localités?(Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN (février 2014) pour obtenir des précisions sur ce terme.) Voir ci-dessus. |
Non |
| Y a-t-il un déclin continu [inféré] de [la superficie, l'étendue ou la qualité] de l'habitat? Aucun déclin de l'habitat n'est évident. |
Non |
| Y a-t-il des fluctuations extrêmes du nombre de populations? Le nombre de sous-populations reproductrices est passé de 1 à 2 depuis 2002, mais cette fluctuation n'est pas considérée comme extrême. |
Non |
| Y a-t-il des fluctuations extrêmes du nombre de localités? (Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN (février 2014) pour obtenir des précisions sur ce terme.) Le nombre de localités avec indice de reproduction est passé de 1 à 2 depuis 2002, mais cette fluctuation n'est pas considérée comme extrême. |
Non |
| Y a-t-il des fluctuations extrêmes de la zone d'occurrence? L'étendue de la zone d'occurrence a augmenté depuis 2002 en raison de l'inclusion de mentions provenant du nord-ouest de l'Ontario; cependant, cette augmentation est considérée comme une expansion de l'aire de répartition et non comme une fluctuation associée au risque de disparition. |
Non |
| Y a-t-il des fluctuations extrêmes de l'indice de zone d'occupation? Voir ci-dessus. |
Non |
| Sujet | Information |
|---|---|
| Population | Inconnu |
| Sous-population du nord-ouest de l'Ontario Il existe peu de données sur les individus matures. La sous-population compte probablement moins de 50 individus matures (mâles et femelles); la reproduction est probable. |
Inconnu |
| Sous-population de l'île Pelée, en Ontario Il existe peu de données sur les individus matures. La sous-population compte probablement moins de 60 individus matures; la reproduction a été confirmée. |
Inconnu |
| Manitoba/parties de l'Ontario/du Québec/du Nouveau-Brunswick Les individus sont exclus parce qu'il s'agirait d'individus immatures qui se seraient déplacés depuis les populations adjacentes des États-Unis et parce que les indices de reproduction ne sont pas assez forts actuellement. |
S.O. |
| Total Estimé < 110 individus matures |
Inconnu |
| La probabilité de disparition de l'espèce à l'état sauvage est d'au moins [20 % sur 20 ans ou 5 générations, ou 10 % sur 100 ans]. L'analyse de viabilité des populations (AVP) n'a pas été réalisée. | S.O. |
Le taux de mortalité au Canada des renards gris s'étant déplacés depuis les États-Unis empêcherait l'établissement de populations reproductrices et l'expansion de la plupart des sous-populations. Parmi les facteurs limitatifs, on compte la prédation par le coyote et la maladie.
| Sujet | Information |
|---|---|
| Situation des populations de l'extérieur La population du Minnesota s'étend vers le nord; les populations du Michigan, de l'État de New York, du Vermont et du New Hampshire sont stables; la population du Maine s'étend vers le nord-ouest. La population de l'Ohio, près de l'île Pelée, est en déclin. |
Dans la plupart des États américains voisins, les populations augmentent. |
| Une immigration a-t-elle été constatée ou est-elle possible? Les individus des sous-populations du nord-ouest de l'Ontario et du Québec sont présumés être soutenus par l'immigration d'individus provenant des États-Unis. L'immigration depuis les États-Unis vers la sous-population de l'île Pelée n'est pas connue, mais elle serait possible en hiver depuis la terre ferme. Les individus observés au Manitoba, au Québec et au Nouveau-Brunswick auraient émigré des États-Unis. |
Oui |
| Des individus immigrants seraient-ils adaptés pour survivre au Canada? On présume que les conditions environnementales sont semblables. |
Oui |
| Y a-t-il suffisamment d'habitat disponible au Canada pour les individus immigrants? | Oui |
| La possibilité d'une immigration depuis des populations externes existe-t-elle? À l'intérieur de la zone d'occurrence, l'immigration vers la sous-population du nord-ouest de l'Ontario est probable; on ne sait pas s'il y a immigration vers la sous-population de l'île Pelée. |
Partiellement |
| Nature délicate de l'information sur l'espèce | - |
| L'information concernant l'espèce est-elle de nature délicate? | Non |
Espèce désignée « préoccupante » en avril 1979. Réexamen du statut : l'espèce a été désignée « menacée » en mai 2002 et en novembre 2015.
Depuis la publication du rapport de situation sur le renard gris par le Comité sur la situation des espèces en péril au Canada (COSEPAC) en 2002, une deuxième sous-population renfermant des individus reproducteurs s'est établie, et la présence de l'espèce a augmenté au Québec et au Nouveau-Brunswick. Le renard gris est toujours présent au Manitoba, mais il ne s'y reproduit pas. Les individus qui ont été signalés au Nouveau-Brunswick, au Québec et au Manitoba sont considérés comme des individus qui se sont déplacés récemment depuis des populations voisines des États-Unis. En raison de l'absence d'indice de reproduction, les individus du Nouveau-Brunswick, du Québec et du Manitoba ne sont pas considérés comme pertinents aux fins de l'évaluation de la situation de l'espèce.
Dans le cadre des évaluations de la situation d'une espèce par le COSEPAC, les rapports sur les connaissances traditionnelles autochtones (CTA) sont préparés par le sous-comité des connaissances traditionnelles autochtones (sous-comité des CTA). Ces rapports préliminaires compilent et résument les CTA facilement accessibles qui s'appliquent aux évaluations de la situation de l'espèce. Un rapport de collecte de CTA peut être rédigé s'il existe d'importantes lacunes dans les connaissances ou des contradictions majeures entre les CTA et d'autres types de connaissances. Le sous-comité des CTA n'a pas relevé de CTA propres au renard gris au Canada.
Le COSEPAC remercie Jennie Pearce d'avoir rédigé le rapport sur la situation du renard gris, aux termes d'un marché conclu avec Environnement Canada. Graham Forbes, coprésident du sous-comité de spécialistes des mammifères terrestres (SCSMT) du COSEPAC, a supervisé les modifications qui ont été apportées au rapport de situation (après l'acceptation de la version provisoire du rapport) selon les commentaires formulés par les provinces concernées, le SCSMT, des membres du COSEPAC et des spécialistes externes.
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) a été créé en 1977, à la suite d'une recommandation faite en 1976 lors de la Conférence fédérale-provinciale sur la faune. Le Comité a été créé pour satisfaire au besoin d'une classification nationale des espèces sauvages en péril qui soit unique et officielle et qui repose sur un fondement scientifique solide. En 1978, le COSEPAC (alors appelé Comité sur le statut des espèces menacées de disparition au Canada) désignait ses premières espèces et produisait sa première liste des espèces en péril au Canada. En vertu de la Loi sur les espèces en péril (LEP) promulguée le 5 juin 2003, le COSEPAC est un comité consultatif qui doit faire en sorte que les espèces continuent d'être évaluées selon un processus scientifique rigoureux et indépendant.
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) évalue la situation, au niveau national, des espèces, des sous-espèces, des variétés ou d'autres unités désignables qui sont considérées comme étant en péril au Canada. Les désignations peuvent être attribuées aux espèces indigènes comprises dans les groupes taxinomiques suivants : mammifères, oiseaux, reptiles, amphibiens, poissons, arthropodes, mollusques, plantes vasculaires, mousses et lichens.
Le COSEPAC est composé de membres de chacun des organismes responsables des espèces sauvages des gouvernements provinciaux et territoriaux, de quatre organismes fédéraux (le Service canadien de la faune, l'Agence Parcs Canada, le ministère des Pêches et des Océans et le Partenariat fédéral d'information sur la biodiversité, lequel est présidé par le Musée canadien de la nature), de trois membres scientifiques non gouvernementaux et des coprésidents des sous-comités de spécialistes des espèces et du sous-comité des connaissances traditionnelles autochtones. Le Comité se réunit au moins une fois par année pour étudier les rapports de situation des espèces candidates.
Remarque : Catégorie « DSIDD » (données insuffisantes pour donner une désignation) jusqu'en 1994, puis « indéterminé » de 1994 à 1999. Définition de la catégorie (DI) révisée en 2006.
Le Service canadien de la faune d'Environnement et Changement climatique Canada assure un appui administratif et financier complet au Secrétariat du COSEPAC.
Noms communs : renard gris (français), Gray Fox (anglais) (Wilson et Reeder, 2005), Grey Fox (anglais canadien)
Noms autochtones : aucun nom n'est utilisé par les Premières Nations dans l'aire de répartition canadienne définie
Classe : Mammifères
Ordre : Carnivores
Famille : Canidés
Genre : Urocyon
Espèce : cinereoargenteus Schreber (1775)
Huit sous-espèces de renard gris sont reconnues en Amérique du Nord (Fritzell et Haroldson, 1982; Wilson et Reeder, 2005), et trois d'entre elles seraient présentes au Canada, le U. c. borealis Merriam, le U. c. cinereoargenteus (Schreber) et le U. c. ocythous Bangs (Banfield, 1974) (figure 2). Sept autres sous-espèces sont reconnues en Amérique centrale et en Amérique du Sud (Fritzell, 1987). La validité des désignations de sous-espèce est abordée dans les sections Structure spatiale et variabilité de la population et Unités désignables.
Le renard gris (figure 1) appartient à la famille des Canidés, qui comprend les chiens, les loups, les renards et les chacals. Sa taille est semblable à celle du renard roux (Vulpes vulpes). Le renard gris possède une longue bande longitudinale foncée, constituée de poils raides aux extrémités noires, sur toute la longueur du dos et de la queue (Fritzell et Haroldson, 1982). Sur les flancs et le dos, le pelage est gris argenté, et le duvet est de couleur chamois. Ce renard possède de la fourrure blanche sur les oreilles, la face, la gorge, la poitrine, le ventre et les pattes arrière, mais le bout de la queue est noir, alors qu'il est blanc chez le renard roux. Le renard gris porte une tache noire distinctive sur le museau en dessous de chaque œil et sur la mâchoire inférieure. Le renard gris a les pattes plus courtes que le renard roux, et son museau est plus court, ses empreintes sont plus rondes, ses coussinets plantaires sont plus larges et ses griffes sont plus recourbées que ceux du renard roux (Banfield, 1974). La longueur corporelle incluant la queue est de 136 cm (plage de 107,5 à 156,8 cm); le poids moyen des mâles est de 4,1 kg, alors que celui des femelles est de 3,9 kg (Banfield, 1974).

Description longue de la figure 1
Photo d'un renard gris en vue latérale, la tête faisant face à l'appareil-photo. Une longue bande longitudinale foncée, constituée de poils raides aux extrémités noires, se trouve sur toute la longueur du dos et de la queue. Sur les flancs et le dos, le pelage est gris argenté, et le duvet est de couleur chamois. On voit de la fourrure blanche sur les oreilles, la face, la gorge, la poitrine, le ventre et les pattes arrière, mais le bout de la queue est noir. On voit une tache noire distinctive sur le museau en dessous de chaque œil et sur la mâchoire inférieure.
Il existe peu de données sur la structure spatiale du renard gris aux États-Unis, et il n'en existe aucune sur la population canadienne de l'espèce. Les désignations des sous-espèces de renard gris ont été établies au début du 20e siècle à la suite de la comparaison des mesures morphométriques (c.-à-d. le crâne, les dents et les mesures corporelles) et du pelage de nouveaux spécimens à ceux du spécimen type connu. Le premier type de spécimen/sous-espèce a été décrit dans les Carolines en 1775 et a été nommé par après Urocyon cinereoargenteus cinereoargenteus (Schreber, 1775; Hall, 1981). La désignation de la sous-espèce ocythous a été fondée sur un spécimen du Wisconsin dont le crâne et le corps étaient plus gros, et dont le pelage était plus jaune (Bangs, 1899), et la désignation de la sous-espèce borealis s'est appuyée sur un spécimen du New Hampshire qui était aussi plus gros, en particulier les dimensions des diverses dents (Merriam, 1903). La définition de l'aire de répartition de chaque sous-espèce a été achevée par Hall (1981), qui a comparé des spécimens de musée et précisé les limites de l'aire de répartition de chaque sous-espèce en reliant les localités des spécimens le long des limites de leur aire de répartition respective.
Selon Hall (1981), le renard gris au Manitoba et dans le nord-ouest de l'Ontario est désigné comme étant le U. c. ocythous, c.-à-d. la sous-espèce qui est présente au Minnesota. Le renard gris observé dans la région du lac Érié serait le U. c. cinereoargenteus, et le renard gris de l'est de l'Ontario, du sud du Québec, et peut-être les mentions provenant du Nouveau-Brunswick, seraient le U. c. borealis (figure 2). Le renard gris qui est présent sur la rive nord du lac Érié en Ontario (et à l'île Pelée) est désigné comme étant le U. c. cinereoargenteus,selon une mention marginale de spécimen provenant du comté d'Oxford (Hall, 1981), et cette population recevrait des individus de la sous-espèce cinereoargenteus provenant des États voisins, à savoir le Michigan, l'Ohio et l'est de l'État de New York.
Description longue de la figure 2
Carte de l'aire de répartition du renard gris dans la partie continentale de l'Amérique du Nord, laquelle montre les localités de 8 des 15 sous-espèces. L'aire de répartition du renard gris est vaste et s'étend depuis la frontière du Canada au Manitoba jusqu'au Québec et, vers le sud, jusque dans l'est et le sud des États-Unis et jusque dans le nord de la Colombie et du Venezuela. Le renard gris est absent du nord-ouest des États-Unis et des Grandes Plaines des États-Unis.
Une seule étude (Bozarth et al., 2011) a porté sur la structure génétique de trois sous-espèces dans l'est de l'Amérique du Nord (à savoir borealis, cinereoargenteus et floridus), dont l'aire de répartition de deux d'entre elles (borealis, cinereoargenteus) s'étend jusqu'au Canada. Aucun échantillon en provenance du Canada n'a été inclus dans l'étude. En se fondant sur un échantillon de 303 individus, Bozarth et al. (2011) n'ont trouvé aucune structure génétique nette des haplotypes du renard gris dans l'est des États-Unis. L'absence de structuration laisse penser que les désignations des sous-espèces pourraient ne pas être valides (Bozarth et al., 2011). Des résultats semblables ont été notés chez d'autres mammifères qui ont survécu dans des refuges durant la glaciation et qui ont étendu par la suite leur aire de répartition vers le nord (p. ex. Wooding et Ward [1997]; Aubry et al. [2009]).
Aucune étude rigoureuse fondée sur des statistiques des différences morphométriques, ou différences génétiques, n'a été menée au sujet du renard gris. Ce type d'étude menée sur d'autres espèces de carnivores à forte capacité de dispersion a fait en sorte que le nombre de sous-espèces nord-américaines a diminué considérablement; par exemple, le renard roux (10 sous-espèces, réduites à 2 clades génétiques, Hall [1981]; Kutschera et al. [2013]), le renard gris (Canis lupus) (de 24 à 5 sous-espèces, Hall [1981]; Nowak [1995]), et le grizzli (Ursus arctos) (de 86 à 7, ou à 2, sous-espèces, Merriam [1918]; Rausch [1963]; Hall [1984]). Le nombre de désignations des sous-espèces de renard gris dans l'est de l'Amérique du Nord pourrait diminuer de façon semblable. Depuis la période glaciaire, il ne semble pas y avoir eu d'obstacle aux déplacements du renard gris dans le centre-est de l'Amérique du Nord, lesquels déplacements auraient isolé les populations et favorisé un caractère génétique unique. Les différences dans la taille corporelle ont constitué le principal critère aux fins de désignation des sous-espèces, mais les différences sont clinales, et les dimensions augmentent du sud vers le nord (Hall, 1981), ce qui est une caractéristique commune aux mammifères de l'Amérique du Nord (p. ex. Ashton et al. [2000]) et qui ne constitue pas une base solide aux fins de désignation des sous-espèces (James, 2010; Ennen et al., 2014).
La seule preuve génétique concernant des différences dans les populations de renards gris est que les échantillons provenant du nord-est des États-Unis possèdent 4 haplotypes, comparativement à 36 haplotypes dans les échantillons provenant des populations plus au sud (Bozarth et al., 2011). Cependant, les chercheurs croient que la différence du nombre d'haplotypes résulterait d'une petite population fondatrice qui s'est étendue vers le nord à partir du refuge, plutôt que d'une profonde divergence génétique créée par l'adaptation aux refuges glaciaires (Bozarth et al., 2011). De plus, la différence génétique ne correspond pas aux limites des sous-espèces; la fréquence des haplotypes dans les échantillons de la sous-espèce borealis provenant du Vermont, du New Hampshire et du Maine était semblable à celle des échantillons de la sous-espèce cinereoargenteus provenant de l'État de New York. Un seul échantillon (provenant du nord de l'Ohio) a été examiné dans les autres États des Grands Lacs, et aucune conclusion n'a donc pu être tirée en ce qui concerne le caractère distinctif de la sous-espèce cinereoargenteus qui gagne le sud-ouest de l'Ontario, ou de la sous-espèce ocythous qui gagne la région de Rainy River - Thunder Bay (ci-après « nord-ouest de l'Ontario ») et le Manitoba. Cependant, étant donné la capacité de dispersion de l'espèce et l'absence d'obstacles à sa dispersion dans l'est de l'Amérique du Nord, il est peu probable que le renard gris de la région des Grands Lacs provienne d'un refuge glaciaire distinct de celui qu'a utilisé le renard gris du sud-est, lequel n'a pas produit de structuration génétique (Bozarth et al., 2011). Il pourrait y avoir eu une profonde divergence génétique chez le renard gris des régions montagneuses de l'ouest des États-Unis, mais rien n'indique que ces individus ont gagné le Canada.
Une seule unité désignable est reconnue, en raison de l'absence de structuration génétique, de la non-correspondance des données génétiques et des données morphométriques utilisées pour définir les sous-espèces, et d'un manque général de rigueur dans les désignations des sous-espèces historiques de renard gris (voir Structure spatiale et variabilité de la population). Les zones renfermant des indices de reproduction sont considérées comme des sous-populations parce qu'elles sont actuellement isolées et très espacées les unes des autres au Canada.
Le renard gris est considéré comme l'espèce la plus basale de la famille des Canidés, ce qui indique l'origine nord-américaine des Canidés il y a 10 millions d'années (Lindblad-Toh et al., 2005). Bien que l'abondance de l'espèce au Canada soit probablement trop faible pour que le renard gris influe sur les écosystèmes, là où il est abondant aux États-Unis, il contribue à la régulation de l'abondance des petits mammifères. Étant donné que le renard gris consomme beaucoup de fruits, il pourrait aussi jouer un rôle dans la dispersion des graines.
Aux États-Unis, le renard gris a une certaine importance économique en tant qu'animal à fourrure, alors que la récolte du renard gris est interdite au Canada.
L'aire de répartition du renard gris est vaste; elle s'étend depuis la frontière du Canada au Manitoba jusqu'au Québec et, vers le sud, jusque dans l'est et le sud des États-Unis et jusque dans le nord de la Colombie et du Venezuela. L'espèce est absente du nord-ouest des États-Unis et des Grandes Plaines des États-Unis (Fritzell et Haroldson [1982]; figure 2).
Au Canada, la présence du renard gris a été signalée en Alberta, au Manitoba, en Ontario, au Québec et au Nouveau-Brunswick (figure 3; tableaux A1 à A7). Les populations reproductrices semblent limitées à deux parties de l'Ontario.

Description longue de la figure 3
Carte indiquant la zone d'occurrence du renard gris au Manitoba, en Ontario, au Québec et au Nouveau-Brunswick, d'après les mentions récentes (effectuées au cours des 20 dernières années). Les symboles indiquent les localités des mentions récentes et des mentions historiques. La superficie totale de la zone d'occurrence du renard gris est de 625 517 km carrés à l'intérieur du territoire canadien.
Un seul individu du renard gris a été piégé non loin du lac Athabasca (Alberta) en janvier 1950, soit à plus de 2 000 km de la population de renards gris la plus proche (Moore, 1952). L'Alberta ne fait pas partie de l'aire de répartition canadienne du renard gris parce que ce spécimen est considéré comme une mention hors de l'aire de répartition, ou comme une espèce errante dans la province (Moore, 1952; Fritzell et Haroldson, 1982; Smith, 1993), voire peut-être comme un spécimen élevé en captivité et qui s'est échappé (Fritzell, 1987).
La première mention confirmée du renard gris au Manitoba est celle d'un mâle adulte piégé en 1957, près de St. Adolphe; il existe une autre mention non confirmée d'un individu à Sprague en 1946-1947 (Sutton, 1958). On compte moins de 10 mentions confirmées et de nombreuses mentions non confirmées, lesquelles proviennent toutes du sud-est du Manitoba, principalement près de Sprague, du parc provincial Whiteshell et de St. Adolphe et à proximité de Hadashville (tableau A1). Le rapport du COSEPAC de 2002 mentionne une communication personnelle d'I. McKay, biologiste provincial spécialiste des animaux à fourrure, selon laquelle six ou sept individus ont été récoltés chaque année (les dates n'ont pas été fournies) dans la région du parc Whiteshell (Judge et Haviernick, 2002). Aucune mention de ces individus n'est disponible, et il existe seulement une mention confirmée provenant du parc Whiteshell, en 2002. Aucune des données de capture d'animaux à fourrure de 1978 à 2014 ne concerne le renard gris (Manitoba Wildlife Branch, 2015). Bien qu'un nombre aussi élevé de captures par le passé soit intéressant, il manque néanmoins des données supplémentaires sur les captures d'individus confirmées et sur la reproduction de ces individus; il n'existe aucune indication de la présence régulière du renard gris dans le parc Whiteshell actuellement (Berezanski, comm. pers., 2014). Le renard gris continue d'être signalé de manière sporadique dans le sud-est du Manitoba, principalement dans des cas de mortalité routière, durant l'hiver (Berezanski, comm. pers., 2014). Les mentions se trouvent toutes à moins de 100 km, et le plus souvent à moins de 60 km, de l'aire de répartition connue de l'espèce au Minnesota. La reproduction n'a pas été confirmée au Manitoba, et les quelques individus qui ont été signalés depuis 2000 sont probablement des individus qui se sont dispersés à partir du Minnesota (Berezanski, comm. pers., 2014). Le gouvernement du Manitoba ne considère pas le renard gris comme une espèce résidente qui se reproduit dans la province.
Bien que des mentions du renard gris datent d'avant l'arrivée des colons européens (voir Fluctuations et tendances), au cours des 50 dernières années, la présence de l'espèce a été signalée dans quatre régions seulement : (1) la région de Rainy River - Thunder Bay (nord-ouest de l'Ontario = nord-ouest du lac Supérieur, le long des États américains); (2) l'île Pelée; (3) la rive nord du lac Érié, de Windsor à Niagara Falls; et (4) la rive nord-est du lac Ontario et du fleuve Saint-Laurent. Même si le renard gris est présent aux environs de Sault Ste. Marie, au Michigan (Rollings, comm. pers., 2014), il n'a pas été signalé dans la ville voisine de Sault Ste. Marie, en Ontario (Siccoly, comm. pers., 2014).
(1) Nord-ouest de l'Ontario (tableau A2)
Le renard gris a été signalé la première fois en 1944 dans le district de Rainy River (Banfield, 1974) et a été observé depuis de Rainy River à Dorion (Ontario); de plus, la présence de 25 à 28 individus a été confirmée depuis 1944 (Judge et Haviernick, 2002; Van den Broeck, 2014a). Les observations semblent avoir augmenté depuis 2006 (Van den Broeck, 2014a). Au début de mars 2014, un mâle mature (4,0 kg) et une femelle mature (4,2 kg) se déplaçant ensemble ont été tués par un véhicule près de Rainy River (Van den Broeck, 2014b). Une autopsie subséquente n'a révélé aucun indice de reproduction (Sadowski, comm. pers., 2015). L'analyse des contenus stomacaux de ce couple d'individus a montré que l'estomac du mâle contenait une petite quantité (25 ml) de pommettes (Malus sp.) et du maïs concassé, alors que l'estomac de la femelle était vide et que la partie inférieure de l'intestin ne contenait que quelques graines de tournesol décortiquées. Les deux individus étaient maigres et avaient peu de graisse corporelle (Sadowski, comm. pers., 2015). De plus, un seul individu a été observé dans la même grande région, en novembre 2013 (Van den Broeck, 2014b). En 2010, deux individus ont été observés ensemble non loin de Thunder Bay et, en 2013, quatre individus ont été observés ensemble dans la même grande région (Van den Broeck, 2014a). À l'été 2015, une femelle en lactation a été observée périodiquement (et photographiée le 1er juillet), dans le secteur du chemin Mountain, au sud de Thunder Bay, et cette observation correspondrait à l'indice le plus solide de l'existence d'une population reproductrice dans la région (Deschamps, comm. pers., 2015).
(2) Île Pelée (tableau A3)
La première mention de renard gris sur l'île Pelée date de 1983, mais on n'est pas certain du moment de l'arrivée de l'espèce sur l'île. La reproduction de l'espèce a été confirmée le 1er juin 1998 lorsqu'un renardeau âgé de 6 semaines a été observé (London Free Press, 1998); à la fin de mai 2011, d'autres observations non confirmées d'individus ayant fait leur tanière sous une maison ont été effectuées (M. McFarlane, données inédites de NCC). Une femelle mature (5,7 kg) tuée par un véhicule et ramassée en décembre 2012 sur l'île Pelée avait des cicatrices utérines, lesquelles indiquaient qu'elle s'était déjà reproduite. Cet individu était en bon état, avec d'importants dépôts de graisse et 178 g de contenus stomacaux, notamment des souris, un petit oiseau chanteur et d'autres aliments non identifiés (Sadowski, comm. pers., 2015). En 2013, des renards gris ont été observés principalement dans le nord de l'île Pelée (Bowman et al., 2013; McFarlane, 2013).
(3) Rive nord du lac Érié (sud-ouest de l'Ontario) (tableau A4)
Compte tenu de la concentration de mentions dans la région de Niagara Falls, le renard gris serait arrivé dans la région depuis l'ouest de l'État de New York en 1948 (Peterson et al., 1953; Banfield, 1974). Entre 1949 et 2002, des individus ont été signalés sur la rive nord du lac Érié, au nord de la péninsule Bruce, et aussi loin vers l'est que Toronto. Un nombre total de 42 à 45 observations confirmées ont été consignées dans la région, mais aucune observation n'a été mentionnée depuis 2002.
(4) Rive nord-est du lac Ontario et du fleuve Saint-Laurent (tableau A5)
La première mention a été celle d'un individu traversant le fleuve Saint-Laurent en 1942 (Banfield, 1974). Peterson et al. (1953) ont cite un communiqué selon lequel le renard gris se reproduisait dans la grande région des comtés de Leeds, de Grenville, de Stormont, de Dundas et de Glengarry en 1952, et selon lequel un ou deux individus étaient abattus chaque année. Un nombre total de 19 observations confirmées ont été signalées entre 1942 et 2014 (Judge et Haviernick, 2002), mais il n'existe aucune mention de reproduction depuis les années 1950. Une mention non confirmée d'un renard de couleur grise qui se trouvait très haut dans un thuya (Thuja sp.) près de Minto a été effectuée en juin 2009 (Linnitt, comm. pers., 2015). Un renard tué par un véhicule a été ramassé sur la promenade des îles du Saint-Laurent en octobre 2008. Bien qu'il ait été impossible de déterminer le sexe de cet individu en raison du mauvais état de la carcasse, il semblait jeune, d'après l'usure de ses dents (Bruce, comm. pers., 2015). Une mention non confirmée d'un renard de couleur grise dont l'extrémité de la queue était noire et qui fréquentait une zone très boisée près du lac Cavendish a été effectuée en septembre 2009 (Arscott, comm. pers., 2015). L'individu a été observé durant plusieurs soirées alors qu'il traversait un petit chemin de gravier situé entre une clairière et le bois. Deux individus ont été observés en 2014, l'un à 28 km au nord-ouest de Smith Falls, et l'autre, à 23 km à l'est de Smith Falls.
Trente-six mentions confirmées de spécimens de renard gris ont été effectuées depuis 1893-1894; elles provenaient des quatre régions suivantes du Québec (MFFP, données inédites; figure 3) : (1) le sud-ouest de la Montérégie; (2) le sud-ouest de l'Estrie; (3) le sud-est de l'Estrie; et (4) le sud-est de la région de Chaudière - Appalaches. Au cours de 20 dernières années (1996-2015), il y a eu 30 mentions de l'espèce.
(1) Sud-ouest de la Montérégie (tableau A6)
Vingt observations confirmées ont été signalées dans le sud-ouest de la Montérégie entre 1975 et 2014, et toutes ces observations ont été effectuées à moins de 42 km de la frontière de l'État de New York. Dans les années 1990, un mâle et une femelle ont été piégés près de Saint-Anicet, à trois semaines d'intervalle (Jolicoeur et al., 2011). Dix des vingt observations ont été signalées au cours des trois dernières années, principalement dans la région de Hemmingford et de Saint-Bernard-de-Lacolle, laquelle se trouve à moins de 6 km de la frontière entre l'État de New York et le Vermont.
(2) Sud-ouest de l'Estrie (tableau A6)
La première mention de renard gris au Canada a été celle d'un individu vu dans le comté de Compton, près de Johnville, durant l'hiver de 1893-1894 (Anderson [1939, cité dans Judge et Haviernick, 2002]). Quatre autres observations ont été mentionnées en hiver, entre 2011 et 2013, notamment une femelle juvénile en 2011, à moins de 45 km de la frontière avec le Vermont et de la frontière avec le New Hampshire.
(3) Sud-est de l'Estrie (tableau A6)
Quatre mentions confirmées ont été signalées dans cette région durant l'hiver. Le premier individu a été piégé dans les années 1940, et les trois autres ont été tués ou piégés entre 2011 et 2013, à moins de 15 km de la frontière du Maine.
(4) Sud-est de la région de Chaudière - Appalaches (tableau A6)
Cinq individus ont été piégés dans la région entre 1986 et 2007, à moins de 45 km de la frontière du Maine.
Le nombre de mentions récentes au Québec, en particulier dans le sud-ouest de la Montérégie et l'ouest de l'Estrie, indique que le renard gris pourrait être résident au Québec, mais les indices de reproduction y sont faibles. Sur les 30 mentions datant des 20 dernières années, 12 individus ont été identifiés à l'âge ou au sexe, à savoir 9 mâles et 3 femelles; sur ce nombre, 4 individus ont été considérés comme des mâles adultes, et 1 individu comme une femelle juvénile (tableau A6). Aucune femelle en lactation ni aucun renardeau n'ont été observés. Toutes les mentions se trouvent à moins de 50 km de populations reproductrices connues ou présumées aux États-Unis. Selon une des hypothèses, le taux de mortalité associée au piégeage accidentel empêcherait l'établissement d'une population reproductrice. L'indice de reproduction le plus fort était une femelle juvénile tuée près de Sherbrooke, le 12 décembre 2011 (tableau A6). Il y aurait eu dispersion en octobre (voir « Dispersion »), et il est possible que cet individu soit né au Vermont et qu'il se soit déplacé jusqu'au Québec dans les quelques mois ayant suivi sa naissance. Selon une autre hypothèse, l'individu pourrait être né au Québec, parce que les femelles se déplacent généralement moins fréquemment que les mâles (voir « Dispersion ») et que les 50 km constituent une distance considérable à franchir. Actuellement, les données probantes indiquent que le renard gris se serait probablement déplacé jusqu'au Québec à partir des États de New York, du Vermont ou du Maine; par conséquent, le renard gris observé au Québec est considéré comme faisant partie de l'aire de répartition canadienne de l'espèce, mais cette mention n'est pas pertinente aux fins de l'évaluation de la situation de l'espèce. Il semble probable qu'il y aura des mentions de reproduction au Québec dans un avenir rapproché.
Deux individus ont été observés au Nouveau-Brunswick (tableau A7). En octobre 2007, un seul mâle juvénile a été piégé près de Rollingdam, dans le comté de Charlotte, au Nouveau-Brunswick (McAlpine et al., 2008). En septembre 2014, un individu a été observé au moyen d'un appareil photo infrarouge activé par le mouvement à l'extrémité nord du lac Oromocto, dans le comté de York, au Nouveau-Brunswick (McAlpine et al., soumis). Les deux individus se seraient déplacés depuis le Maine parce qu'il n'existe aucune autre mention dans la province, et que les mentions les moins lointaines (Bangor, au Maine) sont à une distance de plus de 130 km. Les mentions sont incluses dans le calcul de la zone d'occurrence, mais il est probablement trop tôt pour conclure que le Nouveau-Brunswick fait partie de l'aire de répartition du renard gris. Contrairement au Québec ou au Manitoba, où il existe une population résidente près de la frontière avec les États-Unis, une distance de 100 km sépare la frontière du Nouveau-Brunswick et l'aire de répartition de l'espèce dans le Maine. Il n'existe aucun indice attestant la présence d'une population reproductrice au Nouveau-Brunswick.
Selon les lignes directrices du COSEPAC, une espèce errante est une espèce sauvage dont l'aire de répartition géographique est à l'extérieur du territoire canadien et qui n'a jamais établi une population viable au Canada, mais qui peut être observée de temps à autre au Canada (annexe C, Manuel des opérations et des procédures). Les espèces qui n'ont pas une régularité d'occurrence, comme les espèces errantes, ne sont pas considérées comme valides aux fins de l'évaluation (annexes E3, F9, Manuel des opérations et des procédures). Selon les lignes directrices de l'UICN, le terme « visiteur » s'applique à un taxon qui ne se reproduit pas dans une région, mais qui est aujourd'hui régulièrement présent dans les limites de la région, ou le fut à certaines périodes du siècle passé (IUCN, 2010). Cependant, le concept d'espèce errante ou de « visiteur » ne s'applique pas bien à la situation du renard gris au Canada. Il a été établi que le renard gris était une espèce résidente qui se reproduit dans deux régions du Canada et que l'espèce est admissible à une évaluation; cependant, la majeure partie de l'aire de répartition de l'espèce se trouve dans des régions caractérisées par la présence d'individus apparemment non reproducteurs (immatures) qui se sont déplacés jusqu'au Canada depuis les États-Unis. La reproduction est confinée à l'île Pelée et au nord-ouest de l'Ontario. Les renards gris qui ont été signalés en Alberta, au Manitoba, dans le sud-est de l'Ontario, au Québec et au Nouveau-Brunswick sont considérés comme des individus en dispersion dont la présence est irrégulière ou occasionnelle et qui ne font pas partie d'une population reproductrice établie. Par conséquent, ces mentions sont considérées comme faisant partie de la zone d'occurrence au Canada, mais elles ne sont pas considérées comme ayant une incidence importante sur la viabilité du renard gris au pays et ne sont donc pas incluses dans l'estimation de la population.
La zone d'occurrence et l'indice de zone d'occupation (IZO) sont calculés pour les deux régions où la reproduction du renard gris est probable, ainsi que pour la superficie totale des mentions au Canada (voir Aire de répartition canadienne). Selon les observations des 20 dernières années (c.-à-d. période de 1994 à 2015, valeur arbitraire, mais qui fournit probablement de l'information récente sur les facteurs climatiques et les facteurs de l'habitat qui influent sur la répartition du renard gris - voir Tendances de l'habitat et Fluctuations et tendances), la zone d'occurrence totale de l'espèce est de 625 517 km2, et l'IZO (quadrillage de 2 km × 2 km) est de 200 km2. Selon l'ensemble des mentions récentes (c.-à-d. des 20 dernières années) concernant les deux sous-populations avec indice de reproduction (île Pelée et nord-ouest de l'Ontario), la superficie de la zone d'occurrence est de 35 173 km2, et l'IZO est de 96 km2 (figure 3).
Les données proviennent des centres de données sur la conservation des provinces, des rapports publics sur les observations du renard gris, des statistiques de piégeage et des départements responsables de la faune des États américains. Un relevé systématique à l'aide d'un appareil photo à déclenchement automatique a été mené durant la période de 2012 à 2013 sur l'île Pelée (Bowman et al., 2013). La plupart des autres données sont fondées sur des rapports publics d'observations faites par des bénévoles et sur des cas de mortalité routière et de piégeage accidentel.
Les données sur la taille et les tendances de la population aux États-Unis sont principalement tirées des statistiques de chasse et de piégeage. Au Québec et en Ontario, les chasseurs et les piégeurs qui tuent des renards gris doivent le signaler aux autorités et leur envoyer la carcasse, si demandé; lorsque l'individu est capturé accidentellement et remis en liberté par la suite, il n'est pas nécessaire de signaler la capture. Ailleurs au Canada, les chasseurs et les piégeurs ne sont nullement tenus de signaler la capture accidentelle d'un renard gris.
L'utilisation que fait le renard gris de l'habitat est peu connue au Canada. Des chercheurs du ministère des Richesses naturelles de l'Ontario, à l'unité de recherche sur la rage, ont installé un collier radio sur un individu mâle adulte et ont suivi cet individu par radiotélémétrie d'octobre 1980 à novembre 1981 dans le comté de Lambton (Ontario), une région où on trouve des champs cultivés entrecoupés de boisés de feuillus dominés par l'érable à sucre (Acer saccharum) à l'intérieur d'un réseau routier (Bachmann et Lintack, 1982). Le renard gris mâle a été repéré le plus souvent dans les boisés et s'est déplacé le plus souvent à moins de 100 m de l'orée du bois. Il s'est déplacé seulement à l'occasion dans les zones ouvertes et moins boisées et n'a traversé des routes qu'occasionnellement (Bachmann et Lintack, 1982). L'ensemble des observations de renard gris à l'île Pelée réalisées à l'aide d'un appareil photo ont été faites dans des forêts de feuillus (Bowman et al., 2013; Bowman, comm. pers., 2014). Ce sont les deux seules études portant sur l'utilisation que fait le renard gris de l'habitat au Canada.
L'utilisation de l'habitat par le renard gris qui a été déterminée dans l'aire de répartition du nord-est des États-Unis est probablement semblable à l'utilisation de l'habitat par l'espèce au Canada. Parmi toutes les autres espèces de renards d'Amérique du Nord, c'est le renard gris qui utilise la plus grande proportion de l'habitat boisé, et le couvert de broussailles et d'arbres constitue une importante composante de son habitat (Cypher, 2003). Dans l'est de l'Amérique du Nord, le renard gris préfère un mélange de champs et de boisés, plutôt que de grandes étendues de forêt homogène (Wood et al., 1958). Au Wisconsin, les zones entrecoupées de forêts et de terres agricoles sont considérées comme un habitat de grande qualité (Richards et Hines, 1953; Petersen et al., 1977). En Illinois, le renard gris utilise davantage les vieux champs que ce à quoi on s'attendait, compte tenu de leur disponibilité, et il utilise moins l'habitat agricole et l'habitat de broussailles que ce à quoi on s'attendait, et il utilise les boisés proportionnellement à leur disponibilité (Follmann, 1973). Le renard gris semble utiliser les vieux champs pour s'alimenter (Follmann, 1973; Yearsley et Samuel, 1980), alors qu'il utilise souvent les boisés, en particulier les peuplements denses ou les broussailles, pour s'abriter durant la journée (Follmann, 1973; Yearsley et Samuel, 1980; Hallberg et Trapp, 1984; Haroldson et Fritzell, 1984).
Dans le nord de son aire de répartition aux États-Unis, le renard gris utilise principalement sa tanière pour la mise bas et l'élevage des petits, mais il arrive parfois qu'il l'occupe en d'autres temps pour éviter les prédateurs (Cypher, 2003). Il peut creuser lui-même sa tanière ou modifier une tanière creusée par d'autres espèces (Schmeltz et Whitaker, 1977). Le renard gris peut aussi aménager sa tanière sous des cordes de bois, sous des amas de broussailles, dans des crevasses rocheuses, dans des billes de bois creuses ou des arbres creux, dans des trous sous des arbustes et sous des maisons abandonnées (Latham, 1943; Sullivan, 1956; Trapp et Hallberg, 1975; Fritzell et Haroldson, 1982). Le renard gris a été observé dans une tanière aménagée dans des arbres creux, à une hauteur de 7,6 m (Grinnell et al., 1937) à 9,1 m (Davis, 1960) au-dessus du sol. Dans les forêts de feuillus de l'est des États-Unis, les tanières font généralement face à l'est, au sud-est ou au sud (Richards et Hines, 1953) et sont situées dans des zones broussailleuses ou boisées (Layne et McKeon, 1956a; Nicholson et al., 1985).
Bien que les sites de repos de jour se trouvent généralement dans la végétation dense (Hallberg et Trapp, 1984; Nicholson et al., 1985), certains renards gris ont été vus aussi à l'ombre de blocs rocheux (Trapp, 1978). Au Canada, Bachmann et Lintack (1982) ont trouvé un seul mâle qui utilisait au moins trois sites de repos, qui se trouvaient tous dans des amas de broussailles. À l'exception d'un site dans un champ en jachère, l'ensemble des sites de repos se trouvaient dans des boisés situés à moins de 50 m de la limite du champ.
Le comportement de grimpeur unique au renard gris est résumé dans Fritzell et Haroldson (1982). Le renard gris présente des adaptations qui lui permettent de grimper aux arbres, comme le fait d'être trapu, d'avoir des pattes assez courtes et de longues griffes recourbées et acérées, et une plus grande capacité de faire tourner la patte avant que d'autres Canidés, ce qui peut l'aider à mieux s'agripper lorsqu'il grimpe aux arbres. C'est pour s'alimenter, se reposer et fuir les prédateurs que le renard gris grimpe aux arbres et dans les arbustes. Il peut grimper sur des troncs d'arbres verticaux sans branche à une hauteur pouvant atteindre 18 m en enserrant le tronc à l'aide de ses pattes avant et en poussant son corps vers le haut à l'aide de ses pattes arrière. Il est peu probable que la plupart des cours d'eau constituent des obstacles à la dispersion du renard gris, car ce dernier nage très bien (Cypher, 2003).
De manière générale, la végétation dans le nord-ouest de l'Ontario se caractérise principalement par des forêts de feuillus et des forêts mixtes ou conifériennes utilisées pour la production de bois, à l'exception de la région des plaines d'Agassiz, dans le nord-ouest de la province, qui renferme environ 31 % de terres agricoles mêlées de forêts (Ontario Partners in Flight, 2008a). Au Manitoba, la province voisine, les individus sont présents dans un couvert forestier semblable à celui des plaines d'Agassiz, mais avec une plus grande proportion de prairies et de terres agricoles, mêlées à des forêts. La disponibilité de l'habitat du renard gris dans le nord-ouest de l'Ontario et le sud-est du Manitoba serait stable.
Le couvert forestier occupe environ 20 % de la superficie de l'île Pelée (Essex Region Conservation Authority, 2011). La plupart des parcelles boisées sur l'île Pelée se trouvent à l'intérieur d'aires protégées et, par conséquent, la disponibilité de l'habitat du renard gris dans l'île Pelée serait stable.
Dans le sud de l'Ontario, la perte d'habitat forestier a été importante depuis le début du 19e siècle, car les forêts ont été converties en cultures. De nos jours, la couverture des terres dans le sud-ouest de l'Ontario et sur la rive nord-est du lac Ontario comprend environ 75 % de terres cultivées, 4 % de champs et 14 % de forêts (Ontario Partners in Flight, 2008b). Le couvert forestier se limite principalement à des boisés situés à l'intérieur d'une matrice agricole; la superficie totale du couvert forestier est stable ou pourrait connaître une légère augmentation en raison des activités de reboisement (Ontario Partners in Flight, 2008b). C'est dans la région d'Essex, dans l'extrême sud-ouest de la province que le couvert forestier est le moins important, et c'est dans le nord de l'aire de répartition du renard gris, sur la rive nord du lac Érié et du lac Ontario, qu'il est le plus important. Dans la région d'Essex, environ 77 % du territoire sert à l'agriculture, et les cultures commerciales, les cultures spéciales, les vergers et les cultures en serre constituent les utilisations agricoles les plus courantes. Le reste de la région comprend 19 % de zones urbaines et environ 6 % de forêts. Dans la région de Niagara, on trouve des superficies semblables de couvert arbustif et arboré (Niagara Region Conservation Authority, 2013), et l'abondance la plus grande de couvert forestier se trouve le long de l'escarpement du Niagara ainsi que dans le sud-est de la région (Niagara Region Conservation Authority, 2013). Bien qu'il soit prévu que la disponibilité de l'habitat du renard gris (zones broussailleuses/boisées entrecoupées de champs) demeure stable au fil des années, les paysages fortement agricoles n'offrent pas l'habitat optimal pour l'espèce, et c'est peut-être parce que le couvert de broussailles et d'arbres n'est pas suffisant pour constituer un abri. La région d'Essex étant la partie continentale du Canada la plus proche de l'île Pelée, la rareté du couvert forestier dans cette région limite peut-être les possibilités d'émigration de l'île à la terre ferme.
L'habitat du renard gris dans le sud de l'Estrie, au Québec, consiste principalement en un mélange de zones agricoles, de champs et de forêts. Il existe des forêts jeunes et matures dans une grande partie du sud du Nouveau-Brunswick, où deux individus ont été signalés. Étant donné que le renard gris préfère un mélange de champs et de boisés et qu'il utilise aussi des habitats suburbains, il est probable que l'habitat végétal convenant à l'espèce soit disponible dans le sud du Québec et le sud-ouest du Nouveau-Brunswick.
Très peu de recherches ayant été menées sur le renard gris dans l'ensemble de son aire de répartition, il faudra recueillir d'autres données fondamentales sur l'écologie et la démographie de l'espèce. Les données qui sont résumées dans la présente section sont tirées principalement d'études réalisées aux États-Unis.
Tous les renards d'Amérique du Nord sont des prédateurs opportunistes, et le renard gris est peut-être l'espèce de renard la plus omnivore (Cypher, 2003). Le renard gris mange des mammifères de petite et de moyenne taille, de la charogne, des oiseaux, des insectes comme les Orthoptères, les Coléoptères et les Lépidoptères, des fruits, du maïs, des noix et des graines (Fritzell et Haroldson, 1982; Cypher, 2003). Lors de récentes autopsies menées sur des renards gris tués par des véhicules et ramassés en Ontario, divers aliments ont été trouvés, y compris des pommettes, des graines de tournesol, des souris, un oiseau chanteur et du maïs (Sadowski, comm. pers., 2015).
Le régime alimentaire du renard gris change selon les saisons, les lapins et les petits mammifères constituant la proportion la plus élevée du régime alimentaire en hiver (Hatfield ,1939; Fritzell, 1987), les insectes (particulièrement les Orthoptères) dominant les contenus stomacaux en été, et les fruits et les noix devenant de plus en plus importants dans le régime alimentaire en automne (Pils et Klimstra, 1975). Les matières végétales, comme les fruits, constituent une grande partie du régime alimentaire de l'espèce tout au long de l'année.
Il est difficile de déterminer le rapport des sexes et la structure d'âge dans les populations de renards gris parce que la plupart des estimations sont fondées sur des données de piégeage, lesquelles peuvent présenter un biais en faveur des captures de mâles juvéniles (Fritzell, 1987). Davis et Wood (1959) ont mentionné que les individus de moins d'un an constituaient 48 à 61 % de la récolte de renards gris. Au Wisconsin, 60 à 72 % des femelles capturées durant une période de 3 ans étaient âgées de moins d'un an (Richards et Hine, 1953). En Alabama, 7 % des renards étaient des juvéniles (Sullivan et Haugen, 1956). Au Wisconsin, 66 % des renards gris récoltés étaient des juvéniles (de moins d'un an), 16 % étaient des jeunes de l'année (1,5 an), et 15 % étaient des adultes (Root et Payne, 1985). Dans la seule étude portant sur une population ne faisant pas l'objet d'une récolte et menée sur 50 ans en Caroline du Sud, Weston et Brisbin (2003) ont constaté qu'il existait un biais en faveur des femelles (59 %) et que 56 % des individus étaient des adultes.
Il a été présumé que le renard gris était monogame mais, selon de récentes recherches, il pourrait y avoir plusieurs pères dans jusqu'à 57 % des portées (Weston Glenn et al., 2009). Les individus non appariés repèrent leurs partenaires à l'automne, et la parade nuptiale a lieu au début de l'hiver (Fritzell, 1987). Le comportement du mâle durant l'élevage des petits n'est pas bien compris, mais sa présence non loin de la tanière durant cette période indique qu'il pourrait jouer un rôle dans l'élevage des petits (Chamberlain et Leopold, 2000). Nicholson et al. (1985) ont conclu que c'était principalement les femelles qui fournissaient la nourriture aux petits.
La durée de la période de reproduction est variable dans l'aire de répartition géographique du renard gris, et la reproduction est plus tardive dans les populations du nord que dans les populations du sud (Trapp et Hallberg, 1975; Fritzell et Haroldson, 1982; Fritzell, 1987). Dans l'État de New York et au Wisconsin, le point culminant de la reproduction va de la mi-février à la mi-mars (Sheldon, 1949; Richards et Hine, 1953; Layne et McKeon, 1956b; Jackson, 1961; Root, 1981). La période de gestation est de 53 à 63 jours (Fritzell, 1987). Une portée moyenne de 3,7 petits a été observée dans 8 études au cours desquelles il y a eu dénombrement des embryons in utero chez les femelles gravides ou des cicatrices utérines chez les femelles en lactation. De 1 à 9 fœtus ou cicatrices utérines ont été dénombrés, et l'on en a compté 4 le plus souvent (Fritzell et Haroldson, 1982; Root et Payne, 1985). Le rapport des sexes des fœtus et des nouveau-nés ne diffère pas de manière significative de 1:1 (Layne, 1958). Les juvéniles atteignent la taille/le poids adulte à l'âge d'environ 210 jours (Wood, 1958). Les mâles et les femelles du renard gris atteignent la maturité sexuelle à l'âge de 10 mois environ (Follman, 1978; Root, 1981), et la plupart des individus se reproduisent au cours de leur première année (Root et Payne, 1985).
Aucune information n'est disponible sur le taux de survie du renard gris au Canada, et les données à ce sujet provenant des États-Unis sont généralement fondées sur le taux de mortalité dû au piégeage (Fritzell et Haroldson, 1982; Cypher, 2003). Dans les populations visées par la récolte, il est possible que le taux de mortalité des juvéniles soit élevé. En Floride, Lord (1961) a estimé que 43 à 47 % des renards gris étaient morts durant les sept mois suivant leur naissance et que le taux de mortalité des adultes était de 61 à 64 %. Dans le sud de la Géorgie, 50 % des juvéniles sont morts avant la fin de leur premier été, et 90 % de ces individus n'ont pas survécu à leur premier hiver; le taux de mortalité annuel des adultes était de 50 % (Wood, 1958). Dans une aire protégée où il est interdit de piéger le renard gris, le taux de mortalité annuel des adultes était de 31 % (Weston et Brisbin, 2003). La forte pression exercée par la récolte s'est maintenue dans une grande partie de l'aire de répartition de l'espèce aux États-Unis, ce qui indique que le renard gris est assez productif (Fritzell, 1987; Weston et Brisbin, 2003).
Les données sur la longévité du renard gris proviennent des populations qui sont touchées par un taux de mortalité dû au piégeage. Selon ces données, la plupart des individus ne vivent pas plus que 4 ou 5 ans (Wood, 1958; Lord, 1961). L'âge du seul renard radiopisté en Ontario a été estimé à 4,5 ans (Backmann et Lintack, 1982). La durée estimée d'une génération est de 2 ans, selon la méthode 1 (âge moyen des parents) des lignes directrices de l'Union internationale pour la conservation de la nature (UICN) (IUCN, 2014). Aux fins de ce calcul, les taux de survie des juvéniles et des adultes ont été estimés à 0,31 et 0,43, respectivement; le taux de fécondité a été estimé à 3,7; et l'âge à la première reproduction, à moins de 1 an.
Le renard gris serait une espèce du sud, adaptée à la chaleur (Waters, 1964), et la limite septentrionale de l'aire de répartition de l'espèce peut être définie par sa capacité à tolérer de basses températures (voir Fluctuations et tendances). Le renard gris tolère généralement les humains, et certains individus ont été trouvés aux abords des villes aux États-Unis alors que d'autres ont été signalés non loin de mangeoires d'oiseaux dans le nord-ouest de l'Ontario. Les paysages modifiés par l'humain qui présentent un mélange de forêts et de champs constituent un habitat convenable pour le renard gris (voir Utilisation de l'habitat).
Le renard gris s'alimente principalement durant la nuit ou au crépuscule (Fritzell et Haroldson, 1982) et se repose durant le jour. Il est commun d'observer certains déplacements diurnes entre les lieux de repos (Trapp, 1978). Au cours de ses activités nocturnes, l'individu mâle adulte radiopisté en Ontario a effectué souvent des circuits dans les boisés de son domaine vital (Bachmann et Lintack, 1982). Sur l'île Pelée, presque tous les renards gris détectés au moyen d'appareils photo de surveillance de la faune en 2012-2013 ont été photographiés entre 20 h et 9 h, et la période d'activité maximale se situait entre 22 h et 1 h (McFarlane, données inédites, 2013). Un individu a été photographié durant la journée (à 14 h 35; McFarlane, données inédites).
Au Mississippi, les renards gris se sont déplacés plus rapidement durant la période d'élevage des petits (446 ± 85 m/h) et durant l'hiver (432 ± 83 m/h, moyenne + écart-type) que durant la période de reproduction (344 ± 46 m/h) (n = 37; Chamberlain et Leopold, 2000). Ils se sont déplacés davantage au crépuscule (454 ± 69 m/h) et durant la nuit (443 ± 64 m/h) que durant le jour (265 ± 67 m/h).
Chez le renard gris, l'unité sociale de base comprend un mâle adulte, une femelle adulte et leur progéniture, et ce groupe maintient un domaine vital qui exclut dans une grande mesure les autres groupes semblables. On ne sait pas si l'espèce est territoriale. Le renard gris effectue couramment un marquage odorant au moyen de son urine et de ses fèces, et ce comportement peut servir à marquer les limites de son domaine vital (Cypher, 2003). Le renard gris a aussi utilisé des latrines, dans lesquelles un grand nombre d'individus déposent leurs fèces (Trapp, 1978). Au Mississippi, Chamberlain et Leopold (2000) ont constaté qu'il y avait peu de communication entre des femelles et des mâles voisins (se trouvant à une distance de moins de 150 m), ce qui laisse croire que les femelles et les mâles s'évitent sur une telle distance, et que l'espèce pourrait être territoriale.
Les estimations publiées de la superficie du domaine vital proviennent pour la plupart du sud des États-Unis (Fritzell et Haroldson, 1982; Cypher, 2003). La superficie du domaine vital est petite (p. ex. 0,13 à 0,8 km2) dans les secteurs de grande qualité (Richards et Hine, 1953; Fuller, 1978) à grande (p. ex. 28 à 77 km2) dans les secteurs où la nourriture, l'eau et les abris sont rares (Lord, 1961; Nicholson, 1982); la superficie du domaine vital peut varier selon la saison ou la région géographique, et ce, tant pour les mâles que pour les femelles. Le domaine vital des deux sexes peut augmenter durant l'automne et l'hiver. Durant la période d'élevage des petits, le domaine vital de la femelle peut diminuer de 80 %, alors que celui du mâle peut augmenter (Follman, 1973; Nicholson et al., 1985). Au Mississippi, 37 individus ont été radiopistés durant la période de 1991 à 1997. Bien qu'aucune variation du domaine vital selon le sexe n'ait été observée, il y a eu variation du domaine vital selon la saison (domaine vital plus grand durant la période de reproduction, du 1er février au 31 mai). La superficie du domaine vital (où se font 95 % des déplacements) durant la période de reproduction était de 3,5 km2, et de 1,8 km2 à d'autres moments de l'année. La superficie du centre du domaine vital (où se font 50 % des déplacements) a aussi varié selon la saison; elle était plus grande durant la période de reproduction (0,4 km2) et plus petite durant l'hiver (0,2 km2). Le plus important chevauchement des domaines vitaux était celui de mâles et de femelles (chevauchement de 57 %), et le chevauchement le moins important était celui de mâles voisins (chevauchement de 1 %). Le centre des domaines vitaux semble utilisé exclusivement par des couples résidents (Chamberlain et Leopold, 2000).
À l'âge de 3 mois environ, les petits recherchent de la nourriture en compagnie de leur mère, et ils le font de manière indépendante à l'âge de 4 mois (Nicholson et al., 1985). Des cas de dispersion des jeunes ont été signalés de septembre à décembre (Nicholson et al., 1985), et la plupart des individus qui se sont dispersés l'ont fait à l'automne (Lord, 1961), et probablement en octobre (Nicholson et al., 1985). Les juvéniles sont demeurés dans l'unité familiale jusqu'à l'âge de 8 mois environ (Trapp et Hallberg, 1975; Nicholson et al., 1985). D'une population à l'autre, il y a variation de l'étendue de la dispersion (Cypher, 2003). En Alabama, Nicholson et al. (1985) ont constaté que 100 % des mâles (N = 3) s'étaient dispersés à partir de leur territoire natal, alors qu'aucune femelle (N = 6) ne l'avait fait; dans l'État de New York, 73 % des mâles (N = 11) et 63 % des femelles (N = 8) se sont dispersés (Tullar et Berchielli, 1982).
Des distances de dispersion de 9, 16 et 24 km (Nicholson et al., 1985) et deux distances de dispersion de plus de 24 km (Tullar et Berchielli, 1982) ont été signalées pour des individus mâles du renard gris. Le mâle immature qui a été piégé en octobre au Nouveau-Brunswick se trouvait à 135 km au nord des mentions connues du renard gris dans le Maine (McAlpine et al., 2008).
De manière générale, les mâles se dispersent sur une plus grande distance que les femelles; Tullar et Berchielli (1982) ont signalé une distance de dispersion moyenne de 7 km (plage de 0,3 à 22 km) pour 8 individus femelles, comparativement à une distance de dispersion moyenne de 14 km (plage de 0,0 à 33 km) pour 11 individus mâles. Nicholson et al. (1985) n'ont observé la dispersion que chez les mâles. Cependant, la présence d'individus femelles a été aussi signalée loin de leur territoire natal; Sullivan (1956) a mentionné une distance de dispersion de 83 km, et Sheldon (1953) a retrouvé une femelle, qui avait été marquée, à 84 km de son lieu de naissance, et ce, 3 ans après avoir été marquée au stade de juvénile. Dans l'État de New York, une femelle juvénile s'est déplacée sur une distance de 18 km en 1 mois (Sheldon, 1953). Aucune migration saisonnière n'a été signalée chez le renard gris.
Plusieurs prédateurs chassent le renard gris, notamment l'Aigle royal (Aquila chrysaetos) (Mollhagen et al., 1972), le chien domestique (Tullar et Berchielli, 1982), le lynx roux (Lynx rufus) et le coyote (Canis latrans) (Fedriani et al., 2000; Farias et al., 2005).
Il existe des données montrant que les coyotes ont un impact négatif sur les populations sympatriques de renards gris. L'abondance des coyotes et celle des renards gris étaient inversement liées en Californie (Crooks et Soulé, 1999; Fedriani et al., 2000); les renards gris ont évité les stations de pistage qui ont été fréquentées par le coyote durant la même nuit (Crooks et Soulé, 1999), et le nombre de renards gris a augmenté après l'élimination des coyotes au Texas (Henke et Bryant, 1999). Le coyote tue le renard gris (Wooding, 1984; Farias et al., 2005) et pourrait être un grand prédateur de l'espèce; 7 des 12 renards gris radiopistés retrouvés morts en Californie avaient été tués par des coyotes (Fedriani et al., 2000). L'habitat et la nourriture étant répartis entre les deux espèces et le fait que le renard gris peut grimper aux arbres expliquent peut-être une certaine coexistence entre le coyote et le renard gris (Wooding, 1984; Cypher, 1993), cependant, dans les secteurs où la densité des coyotes est élevée, le coyote aura un impact sur le renard gris (Fedriani et al., 2000).
Aucune donnée ne montre que le renard roux fait concurrence au renard gris ou qu'il en limite l'abondance ou la répartition. En Illinois, le chevauchement des domaines vitaux du renard roux et du renard gris est beaucoup plus important que le chevauchement des domaines vitaux de l'une ou l'autre de ces espèces (Follmann, 1973). Le renard gris digère plus facilement les fruits, il a une gueule plus étroite et des carnassières plus petites que le renard roux. Ces caractéristiques permettent peut-être au renard gris d'être un herbivore plus efficace et un carnivore moins efficace que le renard roux (Jaslow, 1987). Là où les deux espèces coexistent, elles semblent éviter la compétition directe, parce que le renard gris mange principalement des matières végétales et qu'il utilise surtout les zones boisées, alors que le renard roux se nourrit principalement de petits mammifères et qu'il utilise les zones plus ouvertes et cultivées du même paysage (voir Utilisation de l'habitat). Au Maryland, la consommation de plantes selon le poids était de 48 % pour les plantes et de 35 % pour les mammifères chez le renard gris, par comparaison à 10 % pour les plantes et 69 % pour les mammifères chez le renard roux (Hockman et Chapman, 1983).
Il semble peu probable qu'il y ait des hybrides de renard roux et de renard gris, parce que les deux espèces appartiennent à des genres distincts et qu'elles sont probablement incompatibles. Aucune donnée ne montre qu'il y a hybridation, à l'exception d'une mention non confirmée provenant de l'Ohio, laquelle repose sur la description d'une peau vue dix ans plus tôt par un négociant de fourrures (Bezdek, 1944).
Le renard gris est résistant à la gale animale (Sarcoptes scabiei) et à la filaire des chiens (Dirofilaria immitis), deux parasites qui sont fatals ou gravement débilitants pour la plupart des autres Canidés sauvages, comme le renard roux (Stone et al., 1972; Simmons et al., 1980). Les seules maladies qui ont été mentionnées fréquemment chez le renard gris sont la maladie de Carré et la rage (Gier, 1948; Nicholson et Hill, 1984; Davidson et al., 1992). La maladie de Carré et la rage sont presque toujours fatales pour le renard gris (Fritzell, 1987; Yuan Chung Zee, 1999) et peuvent donc limiter les populations de l'espèce lorsque la densité des individus est élevée. Une étude portant sur 157 renards gris malades ou morts entre 1972 et 1989 a révélé que la maladie de Carré était une cause de mortalité plus importante que l'ensemble des autres maladies combinées (Davidson et al., 1992). Aux États-Unis, des réductions des populations locales ont été observées en raison de la maladie de Carré (Nicholson et Hill, 1984) et d'épizooties de rage (Trapp et Hallberg, 1975; Steelman et al., 2000).
Les indices de reproduction sont limités à deux sous-populations (île Pelée [en Ontario] et nord-ouest de l'Ontario). Les renards gris signalés en Alberta, au Manitoba, dans le sud-est de l'Ontario, au Québec et au Nouveau-Brunswick sont considérés comme des individus se dispersant de manière irrégulière ou occasionnelle, qui ne font pas partie d'une population reproductrice établie et qui ne sont pas jugés importants pour la viabilité de l'espèce au Canada. Par conséquent, seules les mentions provenant des deux régions où la reproduction est connue/probable servent aux estimations de la population.
La taille de la population pour l'ensemble des localités est estimée à partir des observations effectuées par le public, des individus tués sur les routes ou des données sur les pelleteries. Aucun relevé systématique visant le renard gris n'a été mené au Manitoba, dans le nord-ouest de l'Ontario, au Québec ou au Nouveau-Brunswick et, dans ces régions, le public n'est pas sollicité activement pour communiquer ses observations. Les observations du renard gris au Manitoba et au Nouveau-Brunswick ne sont pas suivies par les centres de données sur la conservation des provinces parce que la présence de l'espèce dans ces provinces est considérée comme accidentelle. En Ontario et au Québec, les règlements sur la chasse et le piégeage exigent la déclaration de tout renard gris tué (voir Statuts et protection juridiques). Le Centre d'information sur le patrimoine naturel de l'Ontario consigne les observations du renard gris dans la province.
Des appareils photo actionnés par le mouvement et des échantillons de poils prélevés sur des poteaux odorants ont servi au suivi du renard gris à 30 localités sur l'île Pelée, du 31 mai 2012 au 13 mai 2013. À chaque station, on a installé un appareil photo actionné par le mouvement du côté boisé de la lisière d'une forêt/d'un champ, et on a installé une station de frottage à une distance de 5 à 15 m devant l'appareil photo et appâté cette station avec de l'essence de cataire et de la poudre de cataire. Chaque station a fonctionné durant des périodes de 2 à 4 semaines, de nombreuses fois durant l'année. Des échantillons de poils ont été recueillis aux stations de frottage (Anon, 2012).
Les données sur la taille et les tendances de la population aux États-Unis sont principalement fondées sur des données de chasse et de piégeage, même si aucun État n'exige que les trappeurs déclarent leurs prises. Les données sur la récolte étant souvent biaisées en raison de leur dépendance à l'égard de facteurs comme le nombre de trappeurs et de chasseurs, l'effort individuel des trappeurs, les espèces visées (en général, ce sont des espèces autres que le renard gris, comme le coyote) et les prix du marché, elles ne peuvent donc qu'indiquer approximativement les tendances de la population.
Les estimations de la densité de renards gris en Amérique du Nord sont de 1,2 à 2,1 individus par km2 (Trapp et Hallberg, 1975). Cependant, il est difficile d'obtenir de telles estimations parce que les renards ont de grands domaines vitaux, qu'ils sont très discrets et qu'ils ne sont pas faciles à capturer (Cypher, 2003). De plus, la densité des renards est touchée par la qualité de l'habitat, la disponibilité de la nourriture, la compétition interspécifique, les maladies et l'exploitation par les humains. L'estimation de la densité de renards gris peut donc varier grandement à l'échelle spatiale (à l'intérieur des habitats et d'un habitat à l'autre) et à l'échelle temporelle (selon la saison et selon l'année) (Cypher, 2003).
Peu de données sur la dynamique de la population de renards gris sont disponibles (Cypher, 2003). En Pennsylvanie, Richmond (1952) a signalé que le nombre d'individus piégés pour la prime était plus élevé dans les années où la période de janvier à mars était plus humide et plus chaude que la moyenne, et que ce nombre était plus faible dans les années où cette période était plus froide et plus sèche; il a été présumé que les données sur les primes étaient corrélées avec la taille de la population.
La fréquence des rencontres confirmées avec des renards gris dans le nord-ouest de l'Ontario semble avoir augmenté au cours des 10 dernières années, et les observations sont le plus souvent associées à un réseau routier principal (Van den Broeck, 2014a). Après examen des données à l'appui des observations de l'espèce, des individus tués sur les routes et des relevés de piégeage dans le nord-ouest de l'Ontario, Van den Broeck (2014a) a estimé qu'entre 25 et 28 individus confirmés avaient été signalés depuis 1944 et que 21 individus confirmés avaient été signalés au cours des 10 dernières années (voir Aire de répartition canadienne). Les observations d'une femelle en lactation et de nombreux individus se déplaçant en groupes au cours des quelques dernières années impliquent que l'espèce se serait reproduite dans le nord-ouest de l'Ontario. Des observations anecdotiques du renard gris dans le sud-est du Manitoba montrent que l'espèce y est présente, du moins en hiver, mais aucune donnée ne précise s'il existe une population résidente d'individus reproducteurs, et il est peu probable que ces individus fassent partie de la population de Rainy River. Il est probable que la sous-population compte moins de 50 individus matures.
En 2012-2013, le renard gris a été signalé à 16 reprises à 8 localités au moyen d'appareils photo actionnés par le mouvement. Sept de ces localités se trouvaient dans la partie nord de l'île Pelée, et c'est à 4 de ces 7 localités que le renard gris a été le plus fréquemment observé (Bowman et al., 2013). Le nombre d'individus n'a pas été déterminé. En 2001, Judge et Haviernick (2002) ont estimé que la population de renards gris à l'île Pelée comptait environ 60 individus, selon une communication personnelle avec un chasseur et naturaliste de la région (N. Beattie), qui avait vu des tanières de renards gris et qui avait estimé que le nombre de couples reproducteurs était de 12 à 15. Si cette estimation est correcte, la densité aurait été de 1,4 individu/km2.
La sous-population ne fait plus l'objet de piégeage, et il existe des aires protégées où la majeure partie des observations ont été effectuées, ce qui indique que l'estimation précédente de jusqu'à 30 individus matures s'applique encore, et que ce nombre pourrait être plus élevé. Cependant, l'île occupe seulement 42 km2, et le renard gris n'a pas été signalé en plus grande abondance qu'auparavant (tableau A3); il est probable que la sous-population compte moins de 60 individus matures.
Dans la partie continentale voisine du sud-ouest de l'Ontario, la dernière mention confirmée de l'espèce remonte à 2002 (tableau A4). Il est peu probable qu'il y existe davantage que quelques individus, et il est possible que ces individus se soient dispersés à partir des États-Unis. Cette région de l'Ontario (comté d'Essex) a relativement peu de couvert forestier, ce qui limite peut-être la persistance du renard gris dans la région.
Seize mentions confirmées d'individus ont été signalées entre 1942 et 1985 sur la rive nord-est du lac Ontario. Deux mentions d'individus ont été confirmées entre 2000 et 2008, et deux autres mentions d'individus l'ont été en 2014, ce qui indique que le renard gris peut à l'occasion se disperser dans cette région à partir de l'État de New York ou du Québec (tableau A5).
La fréquence des rencontres avec le renard gris confirmées au Québec semble avoir augmenté considérablement au cours des 10 dernières années (Dalpé-Charron, comm. pers.). Il y a eu 34 individus confirmés depuis 1894, et 24 individus confirmés au cours des 10 dernières années (voir Aire de répartition canadienne). Le nombre d'observations au cours des 5 dernières années et la présence d'un mâle et d'une femelle piégés dans la même région indiquent que le renard gris est peut-être une espèce résidente au Québec, bien qu'aucune reproduction n'y ait été confirmée (tableau A6). Il est probable qu'il existe moins de 50 individus matures et que bon nombre d'entre eux sont des individus s'étant dispersés à partir des États-Unis.
Deux individus ont été signalés au Nouveau-Brunswick (voir Aire de répartition canadienne). Ces mentions concernent fort probablement des individus qui se sont dispersés à partir du Maine. Compte tenu de l'expansion vers le nord-est de la population de renards gris du Maine (DePue, comm. pers.), il est possible que d'autres individus s'étant dispersés soient présents au Nouveau-Brunswick, mais qu'ils n'y aient pas été signalés. Cependant, les activités de piégeage sont communes dans le sud-ouest du Nouveau-Brunswick (Cormier, comm. pers., 2014) et, si le renard gris y était présent en grande abondance, le nombre de mentions serait probablement plus grand.
Des données archéologiques (Wintemberg, 1921, 1928, 1936, 1939, 1948; Downing, 1946) indiquent qu'avant l'occupation européenne, le renard gris était presque aussi commun que le renard roux dans le sud de l'Ontario, et aussi loin que Midland (Downing, 1946). Aucun renard gris n'a été mentionné dans les dénombrements effectués par les Européens, et il semble que l'espèce n'était pas présente au Canada avant la colonisation du sud du Québec à la fin du 19e siècle et celle du sud de l'Ontario à la fin des années 1940 (voir Aire de répartition canadienne). Une disparition semblable du territoire, suivie d'une recolonisation, a été mentionnée dans certaines parties du Maine et de la Nouvelle-Angleterre (Waters, 1967). La cause du déclin n'est pas connue, mais on sait que le déclin a concordé avec l'arrivée des Européens et le déboisement et la récolte non durable de nombreuses espèces animales sauvages qui ont suivi (Bozarth et al., 2011). Cependant, Downing (1946) a présumé que le renard roux était rare ou absent en Ontario au moment de la colonisation parce que l'espèce n'a pas été mentionnée par les auteurs du temps des colonies et parce qu'une espèce animale de grande taille, capable de grimper aux arbres, aurait sans doute été remarquée si elle avait été aussi abondante que les données archéologiques ne le laissent croire.
La disparition de l'espèce du territoire pourrait être liée aux changements climatiques; Waters (1964) a suggéré que l'aire de répartition du renard gris s'était élargie vers le nord et vers l'est durant la période de chaleur appelée hypsithermal (de 5 000 à 2 000 ans av. J.-C.; Dorf, 1959) et qu'elle s'était ensuite contractée vers le sud lorsque la région a connu un refroidissement. Le processus s'est répété durant l'anomalie climatique médiévale (de 800 à 1 300 apr. J.-C.), période au cours de laquelle le renard gris s'est déplacé vers le nord-est; l'aire de répartition de l'espèce s'est par la suite rétractée durant le Petit Âge glaciaire (de 1 500 à 1 850) (Bozarth et al., 2011). Le mécanisme responsable de l'évolution de la population n'est pas connu mais, selon Root et Payne (1985), la portée moyenne comptant un petit nombre de petits observée dans le cadre de leur étude menée au Wisconsin pourrait indiquer une tolérance moins grande aux conditions environnementales et nutritionnelles ainsi qu'une sensibilité aux températures froides. Le coût énergétique associé à la vie dans des régions où la neige est profonde pourrait aussi constituer un facteur limitatif (Judge et Haviernick, 2002).
Dans les années 1930, l'aire de répartition du renard gris a pris de l'expansion vers le nord à partir du centre de l'est des États-Unis (Hamilton, 1943), peut-être en raison du réchauffement climatique et/ou de l'augmentation de la quantité de jeunes forêts sur les terres agricoles abandonnées (Palmer, 1956; Bozarth et al., 2011). L'arrivée du renard gris dans le nord-ouest de l'Ontario et au Manitoba dans les années 1940 aurait été liée à l'expansion vers le nord de l'aire de répartition des lapins (Sylvilagus spp.) (Sutton, 1958). L'expansion vers le nord de l'aire de répartition du renard gris au Wisconsin et au Minnesota serait associée aux changements climatiques (Judge et Haviernick, 2002), mais pourrait aussi être associée à des changements dans le couvert forestier et à des périodes intenses de récolte de bois (Erb, comm. pers., 2014). Dans les années 1940, le renard gris était absent du Maine (Hamilton, 1943), mais, l'espèce est assez commune de nos jours dans la moitié sud de l'État; en moyenne, 134 individus par année ont été piégés de 2001 à 2006 (McAlpine et al., 2008).
L'aire de répartition canadienne du renard gris se trouve le long de la frontière sud du centre-est du pays et correspond à la limite septentrionale de l'aire de répartition de l'espèce en Amérique du Nord. Au Canada, les mentions ont été trop peu nombreuses pour pouvoir déceler des tendances récentes, mais il est probable que toute tendance de la population au Canada est fortement déterminée par l'expansion et la diminution de l'aire de répartition des populations dans des États américains voisins.
Le renard gris s'est dispersé vers le Canada à partir des populations des États-Unis à compter du début du 20e siècle (Downing, 1946) et il continue sans doute à le faire. La présence de l'espèce au Canada après une absence d'au plus 350 ans est due entièrement à l'immigration d'individus en provenance des États-Unis (Downing, 1946). Si la population canadienne de renards gris venait à disparaître encore, il se pourrait que des individus en provenance du nord des États-Unis finissent par repeupler l'aire de répartition canadienne de l'espèce.
Au Minnesota, l'aire de répartition du renard gris semble prendre de l'expansion (Erb, comm. pers., 2014). Avant les années 1980, l'espèce était généralement limitée aux forêts de feuillus du sud au centre du Minnesota et jusqu'au nord-ouest de l'État (MNDNR, 2014). L'espèce est rare dans les comtés du nord-ouest des États-Unis qui sont voisins du Manitoba et où l'agriculture est très intensive (Erb, comm. pers., 2014). Cependant, le renard gris a été observé plus fréquemment au cours des 10 dernières années dans le nord-est du Minnesota, État voisin de l'Ontario, et sa présence a été mentionnée pour la première fois dans la région dans les années 1980 (Latimer, 2014). L'expansion de la population de renards gris dans les comtés du nord-est du Minnesota explique sans doute l'augmentation des observations de l'espèce dans le nord-ouest de l'Ontario au cours des 10 dernières années.
Le renard gris est présent dans l'ensemble du Michigan, et ses populations y sont stables (Rollings, comm. pers., 2014). L'espèce est signalée aussi dans l'ensemble de l'Ohio, mais à une densité faible dans certaines zones à proximité du lac Érié (OHDNR, 2014a). Bien que la tendance des populations à l'échelle de l'Ohio ait beaucoup fluctué, la tendance générale est au déclin à long terme (OHDNR, 2014a). Ce déclin est indiqué par le nombre plus élevé d'observations/1 000 heures au début des années 1990; il est possible que la population se soit stabilisée au cours des dix dernières années (Prange, comm. pers., 2014). Il existe une population qui est stable le long de la frontière entre l'État de New York vers l'Ontario (Schiavone, comm. pers., 2014). L'immigration d'individus à partir de l'État de New York vers l'Ontario a été signalée dans la région de Niagara Falls (voir Aire de répartition canadienne).
On ne sait pas si des individus en provenance de l'Ohio, du Michigan ou de l'État de New York migrent à l'île Pelée ni, le cas échéant, à quelle fréquence ils le font. Des individus pourraient atteindre l'île en traversant sur la glace durant l'hiver. L'Ohio constituerait la source la plus proche de renards gris pour l'île Pelée, car l'État est situé à 10 km des autres îles et à 12 km de la terre ferme, alors que 15 km séparent l'île Pelée de l'extrémité du parc national du Canada de la Pointe-Pelée, en Ontario. De 1972 à 2011, la zone entre l'Ohio et l'île Pelée a été couverte de glace à plus de 90 % chaque année, à l'exception d'une année (NOAA, 2015; Wong et al., 2012). La glace se forme généralement de la fin de décembre à la fin de mars, et la couverture de glace est maximale de la mi-janvier à la mi-février (Assell, 2004). En 1970, un naturaliste qui est venu dans la région s'est fait dire que les caches de chasse qu'il avait vues sur la rive aux pointes Lighthouse et Fish y étaient installées chaque hiver pour tuer les coyotes et les renards susceptibles d'arriver à l'île et d'y avoir un impact sur la chasse annuelle au faisan de Colchide (Phasianus colchicus) (Pratt, comm. pers., 2015); il est donc probable que le renard gris gagne l'île en traversant sur la glace. Cependant, on ne sait pas si une immigration d'individus provenant de l'Ohio est possible pour la sous-population actuelle de renards gris à l'île Pelée parce que les populations de renards gris en Ohio connaissent des déclins depuis les années 1990 et que l'abondance relative est la plus faible dans les régions qui sont les plus proches du Canada (Ohio DNR, 2015). La fréquence d'un couvert de glace continu entre l'Ohio et l'Ontario au cours des prochaines années ne peut pas être déterminée.
Des déplacements de renards gris à partir de l'État de New York vers le sud-est de l'Ontario ont été signalés pour la première fois en 1942 (voir Aire de répartition canadienne), et des individus en provenance de l'État de New York, du Vermont, du New Hampshire et du Maine gagnent actuellement le Québec. Le renard gris est présent dans l'ensemble du Vermont, mais c'est dans le sud de l'État qu'on le trouve en plus grand nombre. Les données de piégeage indiquent que les captures par unité d'effort ont augmenté au cours des dix dernières années, ce qui indique que la population a peut-être augmenté, bien que rien ne montre qu'elle prend de l'expansion vers le nord (Bernier, comm. pers., 2014). Dans les unités de gestion de la faune du Vermont qui bordent le Québec, un nombre total de 15 individus ont été récoltés depuis 1990, et sept d'entre eux l'ont été depuis 2010 (Bernier, comm. pers., 2014).
Selon les données de piégeage, le renard gris est présent dans l'ensemble du New Hampshire, et c'est dans le nord de l'État que l'effort de capture et le taux de capture sont les plus faibles. Aucune donnée n'indique une augmentation de la population ni une expansion de l'aire de répartition de l'espèce au New Hampshire (Tate, comm. pers., 2014).
L'aire de répartition du renard gris a connu une expansion dans le nord-est du Maine au cours des 20 dernières années (McAlpine et al., 2008). L'aire de répartition actuelle de l'espèce s'étend aussi loin à l'est que la rivière Penobscot, et certains individus pionniers ont été signalés à l'est de cette région, vers le Nouveau-Brunswick (figure 2; DePue, comm. pers., 2014). Il est possible que les observations du renard gris au Nouveau-Brunswick augmentent, parce que la population du Maine étend son aire de répartition vers cette province. Dans le Maine, la limite septentrionale de l'aire de répartition de l'espèce se trouve dans la région de Madrid, qui est située à 90 km environ de la frontière du Québec.
L'expansion de l'aire de répartition du renard gris est probablement limitée par les grandes densités de coyotes (voir Relations interspécifiques), la destruction du couvert forestier (voir Utilisation de l'habitat) et peut-être la neige profonde (voir Fluctuations et tendances). Cependant, il n'existe aucune donnée à l'échelle de la population sur la mesure dans laquelle ces facteurs touchent le renard gris au Canada ou aux États-Unis. Une grande partie du couvert forestier a été détruite au 19e siècle, et il serait difficile d'en quantifier les impacts sur les populations récentes. Des hivers plus doux dans le futur le long de la frontière, du Manitoba au Nouveau-Brunswick, favoriseront sans doute l'expansion du renard gris au Canada, mais la neige profonde prévue au Nouveau-Brunswick pourrait constituer un facteur limitatif pour l'espèce dans la région.
Le calculateur des menaces de l'UICN permet de résumer les menaces connues ou prévues pesant sur le renard gris au Canada (annexe B). L'accent est mis sur les deux sous-populations où la reproduction est confirmée ou probable et où les menaces pesant sur ces sous-populations sont moins évidentes. Dans d'autres parties du pays, les individus qui ne semblent pas se reproduire sont touchés par la mortalité due au piégeage. Or, il est difficile d'appliquer ce scénario au calculateur des menaces; la mortalité causée par les humains n'est pas considérée comme un facteur limitatif, et les menaces sont centrées sur la cohorte d'individus matures (reproducteurs). Dans le présent rapport, le piégeage a été considéré comme une menace, d'une part parce qu'il pourrait avoir un impact sur les individus matures et, d'autre part parce qu'il empêcherait l'établissement d'un plus grand nombre d'individus reproducteurs. L'impact global des menaces est élevé en raison de l'impact de la mortalité due au piégeage dans une grande partie de la zone d'occurrence de l'espèce. Les catégories suivantes sont considérées comme importantes pour la viabilité du renard gris au Canada.
Chasse et piégeage (UICN 5.1)
Les mentions de renard gris au Canada proviennent de la zone située à proximité de la frontière canado-américaine, et il semble que la population de l'espèce aux États-Unis prend de l'expansion vers le nord (voir Répartition). Quelques individus montrent des signes de reproduction et, lorsque leur sexe est connu, la plupart des mentions concernent des mâles, ce qui indique qu'il s'agit d'individus qui se sont dispersés et qui ont émigré des États-Unis. À l'exception des mentions à l'île Pelée, un grand nombre des mentions au Canada concernent des individus tués de manière accidentelle par des chasseurs d'animaux à fourrure. Au Québec, 24 des 30 (80 %) mentions de renard gris des 20 dernières années portaient sur des individus qui avaient été piégés accidentellement (MFFP, données inédites; tableau A6). Cinq des 17 observations de renard gris dans le nord-ouest de l'Ontario réalisées depuis 1944 (Van den Broeck, 2014a; tableau A2) et 1 des 2 mentions provenant du Nouveau-Brunswick (tableau A7) ont été obtenues en raison de prises accidentelles durant le piégeage. Bien que les populations de renards gris puissent résister à une forte pression de récolte (voir Cycle vital et reproduction), les prises accidentelles dans les populations de faible densité au Canada limitent probablement l'établissement de populations reproductrices de renards gris dans certaines parties du pays.
Judge et Havienick (2002) ont mentionné qu'environ six ou sept renards gris étaient récoltés chaque année dans la région du parc provincial Whiteshell dans le sud-est du Manitoba. Cependant, aucune peau n'a été vendue récemment au Manitoba (Berezanski, comm. pers., 2014). Au Québec, le renard gris figure sur la liste des animaux à fourrure et les gestionnaires de la faune peuvent donc adopter rapidement des règlements, si nécessaire. La réglementation du Québec stipule que tout renard gris capturé vivant doit être remis en liberté et que tout individu mort doit être signalé à un agent de conservation de la faune (MFFP, 2014).
La principale cause de mortalité du renard gris aux États-Unis est la récolte par les humains lors du piégeage et de la chasse (Cypher, 2003). Des renards gris sont récoltés dans l'ensemble des États voisins des Grands Lacs et du nord-est des États-Unis ( tableau 1 ). La saison de récolte va du 1er septembre au 31 mars au New Hampshire et du 3 novembre au 31 décembre dans le Maine. Il n'existe aucune limite concernant le nombre de prises. On ne peut déterminer la mesure dans laquelle le nombre de prises aux États-Unis a une incidence sur l'immigration de source externe vers le Canada pour le renard gris.
La remise d'une prime pour le renard gris à l'île Pelée a existé jusque dans les années 1980, après quoi elle a été supprimée parce que ce n'était pas conforme à la Loi sur la chasse et la pêche de l'Ontario (Judge et Haviernick, 2002). N. Beattie, chasseur et naturaliste résident, est cité dans le rapport du COSEPAC de 2002; il précise que les chasseurs de l'île Pelée ont abattu entre six et dix renards gris chaque hiver dans les années 1990 et que rien n'indique que cette pratique a eu une incidence négative sur la population (Judge et Haviernick, 2002). On ne dispose d'aucune donnée récente sur l'existence, ou l'importance, de la récolte illégale ou accidentelle du renard gris à l'île Pelée.
| État | Saison de chasse | Saison de piégeage | Limite de prises | Source |
|---|---|---|---|---|
| Wisconsin | Du 19 octobre au 14 février | Du 19 octobre au 14 février | Aucune | WIDNR (2013) |
| Michigan | Du 15 octobre au 1er mars | Du 15 octobre au 1er mars | Aucune | MIDNR (2013) |
| Ohio | Du 10 novembre au 31 janvier | Du 10 novembre au 31 janvier | Aucune | OHDNR (2014b) |
| New York | Du 25 octobre au 15 février | Du 25 octobre au 10 décembre Note du tableaua Du 25 octobre au 15 février Note du tableaub |
Aucune | NYDEC (2014) |
| Vermont | Du 25 octobre au 8 février | Du 25 octobre au 31 décembre | Aucune | VTDFW (2013) |
| New Hampshire | Du 1er septembre au 31 mars | Du 15 octobre au 1er mars | Aucune | NHFGD (2014) |
| Maine | Du 20 octobre au 28 février | Du 3 novembre au 31 décembre | Aucune | MEDIFW (2013) |
Développement résidentiel et commercial (UICN 1.0)
Une étude menée dans le sud de la Californie a révélé l'existence d'une corrélation négative entre le renard gris et l'intensification de l'urbanisation, et la proximité des zones urbaines (Ordenana et al., 2010.) Cependant, cette étude a été réalisée dans une zone de forte densité humaine (19 millions de personnes), et il est peu probable que l'impact de l'urbanisation se fasse sentir dans l'aire de répartition canadienne de l'espèce.
Routes (UICN 4.1)
La mortalité routière est considérée comme un facteur limitatif; son importance comme menace n'est pas clairement établie.
Le renard gris est vulnérable à la mortalité routière en raison de la grandeur de son domaine vital, de ses grandes distances de dispersion et de son association avec les paysages ruraux. Il n'existe aucune donnée sur la mortalité routière de l'espèce au Canada. Cependant, une étude menée en Louisiane dans le cadre de laquelle des renards gris ont été radiopistés a révélé que 8 des 17 individus morts avaient été tués lors de collisions avec des véhicules. Les chercheurs ont estimé à 34 % la probabilité que le renard gris soit exposé à une mortalité attribuable aux activités humaines (mortalité routière et piégeage combinés).
Au Québec, 3 des 34 observations de renard gris sont liées à la mortalité routière (MFFP, données inédites). Dans l'est de l'Ontario, au moins trois observations de renard gris sont liées à la mortalité routière. Il existe des mentions anecdotiques de renard gris tués sur les routes du sud-est du Manitoba (voir Répartition).
Dans les deux sous-populations où il y a reproduction, 3 des 25 observations de renard gris ont été associées à la mortalité routière dans la sous-population du nord-ouest de l'Ontario, et 2 des 17 observations ont été associées à la mortalité routière dans la sous-population de l'île Pelée. Bien qu'assez peu de véhicules circulent à l'île Pelée, la mortalité routière y constitue néanmoins une cause de mortalité.
Maladies non indigènes envahissantes (UICN 8.1)
Les maladies non indigènes comme la maladie de Carré et la rage sont fatales pour le renard gris et elles ont été signalées chez un grand nombre d'individus sauvages aux États-Unis (Davidson et al., 1992). Lors d'épizooties, ces maladies pourraient constituer d'importants facteurs limitatifs pour l'espèce. En Ontario, deux renards gris ont eu la rage; les deux cas ont été signalés en 1986, laquelle constitue une année record avec le plus grand nombre de cas de rage signalés à ce jour pour l'ensemble des espèces en Ontario (Bachmann, comm. pers., 2014). On ne connaît pas la prévalence de toute maladie chez le renard gris au Canada, parce l'évaluation n'a porté que sur un faible nombre d'individus.
Le nombre de localités de renard gris est inconnu, mais il est sans doute supérieur à 10, ce qui correspond au seuil qui permet d'utiliser le nombre de localités dans une évaluation de la situation d'une espèce. La population reproductrice à l'île Pelée occupe une petite superficie (42 km2), et les menaces sont semblables dans l'ensemble de la sous-population, ce qui indique que cette sous-population compte pour une seule localité. Cependant, la sous-population du nord-ouest de l'Ontario occupe une plus grande superficie (environ 35 000 km2), et l'effort de piégeage varie probablement assez pour que de nombreuses localités existent. Il convient de noter que le concept de localité s'applique difficilement à une espèce mobile dont la répartition est fondée sur des mentions ponctuelles, éparses et rares; il est ardu d'associer ces mentions à une zone particulière du point de vue écologique et géographique pour déterminer une menace spécifique.
En 2015, le renard gris a été désigné espèce menacée par le COSEPAC (et il avait déjà été désigné espèce menacée en 2002). L'espèce figure actuellement parmi les espèces menacées à l'annexe 1 de la Loi sur les espèces en péril (LEP). La Loi sur les parcs nationaux du Canada protège le renard gris et son habitat dans les parcs nationaux (p. ex. parc national du Canada de la Pointe-Pelée, parc national du Canada des Îles-du-Saint-Laurent et site historique de la voie navigable Trent - Severn).
En Ontario, le renard gris a été désigné espèce menacée en 2008 en vertu de la Loi de 2007 sur les espèces en voie de disparition de l'Ontario, et l'espèce et son habitat sont protégés dans cette province. Au Québec, le renard gris n'est pas désigné espèce menacée, vulnérable ou susceptible d'être menacée ou vulnérable en vertu de la Loi sur les espèces menacées ou vulnérables. Tant en Ontario qu'au Québec, les lignes directrices pour la chasse et le piégeage exigent que les renards gris capturés de manière accidentelle soient remis en liberté lorsqu'ils sont vivants, ou qu'ils soient déclarés (et peut-être remis) aux agents de conservation lorsqu'ils sont morts (MRNF, 2014a,b; Loi de 1997 sur la protection du poisson et de la faune de l'Ontario). Aux termes des Forest Management Guidelines for Conserving Biodiversity at the Stand and Site Scales de l'Ontario (OMNR, 2010), les tanières de renard gris qui ont déjà été occupées sont protégées, de même que les tanières qui sont découvertes durant les opérations de gestion forestière.
Au Manitoba et au Nouveau-Brunswick, les mentions de renard gris sont considérées comme accidentelles, et l'espèce n'est pas protégée en vertu de la Loi sur les espèces en voie de disparition de 2007 du Manitoba ni en vertu de la Loi sur les espèces en péril du Nouveau-Brunswick. Le renard gris n'est pas considéré comme un animal à fourrure en vertu de la Loi sur la conservation de la faune du Manitoba. Au Nouveau-Brunswick, les règlements visant la récolte du renard gris seraient les mêmes que ceux qui visent le renard roux parce que le renard gris n'est pas considéré comme présent dans la province et parce que la Loi sur le poisson et la faune du Nouveau-Brunswick ne fait référence qu'au « renard ».
À l'échelle mondiale, le renard gris représente une préoccupation mineure (Least concern) pour l'Union internationale pour la conservation de la nature (UICN) parce qu'il est très répandu et que rien n'indique que ses populations diminuent dans l'une ou l'autre partie de son aire de répartition (Cypher et al., 2008). L'espèce est classée par NatureServe (2014) comme non en péril à l'échelle mondiale (G5) et aux États-Unis (N5). Dans les États voisins des occurrences de l'espèce au Canada, le renard gris est classé comme non en péril (S5 : Maine, Vermont, État de New York, Pennsylvanie), apparemment non en péril à non en péril (S4S5 : New Hampshire, Wisconsin), apparemment non en péril (S4 : Michigan) ou non classé (SNR : Ohio, Minnesota). Les autres cotes attribuées par les États sont présentées au tableau 2 .
Au Canada et en Ontario, NatureServe classe le renard gris dans la catégorie des espèces gravement en péril (N1, S1), et le rapport Les espèces sauvages classe l'espèce dans la catégorie en péril (cote 1; CESCC, 2006, 2011). La présence de l'espèce est considérée comme accidentelle en Alberta, au Manitoba et au Nouveau-Brunswick (CESCC, 2011), et le renard gris est donc non classé par NatureServe et le rapport Les espèces sauvages. Au Québec, la présence du renard gris est considérée comme occasionnelle, et l'espèce est cotée SNA par NatureServe (Gauthier, comm. pers., 2014; MFFP, 2014c).
| Cote de NatureServe | Description | État |
|---|---|---|
| S2 | En péril | Wyoming |
| S3 | Vulnérable | District de Columbia, Iowa, Kansas |
| S3S4 | Vulnérable - apparemment non en péril | Utah |
| S4 | Apparemment non en péril | Colorado, Indiana, Kentucky, Michigan, Missouri, Nebraska, Oklahoma, Oregon |
| S4S5 | - | Louisiane, New Hampshire, Wisconsin |
| S5 | Non en péril | Alabama, Arizona, Arkansas, Caroline du Nord, Connecticut, Dakota du Sud, Delaware, Géorgie, Illinois, Maine, Maryland, Massachusetts, Mississippi, Navajo Nation, Nevada, New Jersey, Nouveau-Mexique, New York, Pennsylvanie, Rhode Island, Tennessee, Texas, Vermont, Virginie, Virginie-Occidentale |
| SU | À l'étude | Dakota du Nord |
| SNR | Non classée | Californie, Caroline du Sud, Floride, Minnesota, Ohio |
Les mentions de renard gris au Manitoba et dans le nord-ouest de l'Ontario ont été effectuées principalement sur des terres privées et des terres de la Couronne gérées à des fins de production de bois d'œuvre. Au Manitoba, la présence de l'espèce a été signalée dans le parc provincial Whiteshell. Aucune mesure de protection particulière de l'habitat du renard gris n'a été établie.
À l'île Pelée, l'habitat du renard gris se trouve dans la Réserve naturelle provinciale de la Pointe Fish, qui est gérée par Parcs Ontario en vertu de la Loi sur les parcs provinciaux et les réserves de conservation, et dans l'alvar du chemin Stone, qui est géré par l'Office de protection de la nature de la région d'Essex conformément à la Loi sur les offices de protection de la nature.L'habitat du renard grisest protégé aussi dans plus de 400 ha de terrains privés qui sont gérés à des fins de conservation par Ontario Nature et Conservation de la nature Canada.
Les mentions de renard gris proviennent principalement des terres privées ou des terres de la Couronne. Il existe des mentions historiques (non confirmées) de l'espèce dans le parc national du Canada des Îles-du-Saint-Laurent (en Ontario) - et un individu a été tué dans ce parc en 2008 (Parks Canada, 2014) -, dans le parc national du Canada de Pointe-Pelée et dans le lieu historique national de la voie navigable Trent - Severn; cependant, aucune mention récente n'a été effectuée dans ces localités (Nantel, comm. pers., 2014).
Dean Berezanski, Provincial Furbearer Biologist, Conservation et Gestion des ressources hydriques Manitoba, Winnipeg (Manitoba)
Chris Bernier, Furbearer Biologist, Vermont Fish and Wildlife, Springfield (Vermont)
Sean Blaney, botaniste et directeur exécutif, Centre de données sur la conservation du Canada atlantique, Sackville (Nouveau-Brunswick)
Jeff Bowman, chercheur scientifique, section de recherche-surveillance en matière de faune, ministère des Richesses naturelles et des Forêts de l'Ontario, Peterborough (Ontario)
Jennifer Boyko, biologiste, gestion des ressources, district de Dryden, ministère des Richesses naturelles de l'Ontario, Dryden
Alain Branchaud, biologiste, rétablissement des espèces en péril, Environnement Canada, Montréal (Québec)
Vivian Brownell, biologiste principale des espèces en péril, Direction des espèces en péril, ministère des Richesses naturelles de l'Ontario, Peterborough (Ontario)
Emmanuel Dalpé-Charron, biologiste et coordonnateur provincial, Ministère des Forêts, de la Faune et des Parcs du Québec, Québec (Québec)
Michael Deschamps, biologiste de secteur, secteur de Thunder Bay Sud, ministère des Richesses naturelles de l'Ontario, Thunder Bay (Ontario)
John DePue, Furbearer/Small Mammal Biologist, Maine Inland Fisheries and Wildlife, Bangor (Maine)
Dave Duncan, gestionnaire, section de la conservation des populations, Service canadien de la faune, Environnement Canada, Edmonton (Alberta)
John Erb, Furbearer Research Biologist, Minnesota Department of Natural Resources, Grand Rapids (Minnesota)
Chris Friesen, gestionnaire de l'information sur la biodiversité, Centre de données sur la conservation du Manitoba, Winnipeg (Manitoba)
Martina Furrer, biologiste, information sur la biodiversité, Centre d'information sur le patrimoine naturel, ministère des Richesses naturelles de l'Ontario, Peterborough (Ontario)
Isabelle Gauthier, coordonnatrice provinciale, espèces fauniques menacées et vulnérables, ministère des Forêts, de la Faune et des Parcs du Québec (Québec)
Neil Jones, chargé de projets scientifiques et coordonnateur des CTA, Secrétariat du COSEPAC, Service canadien de la faune, Environnement Canada, Gatineau (Québec)
Kimberley McNaughton, biologiste, gestion des ressources, district de Nipigon, ministère des Richesses naturelles de l'Ontario
Don McAlpine, directeur, Département des sciences naturelles, Musée du Nouveau-Brunswick, Saint John (Nouveau-Brunswick)
Christy McDonald, analyste des politiques de la faune, section des politiques de la faune, ministère des Richesses naturelles de l'Ontario, Peterborough
Mhairi McFarlane, biologiste de la conservation, sud-ouest de l'Ontario, Conservation de la nature Canada, London (Ontario)
Randall Mooi, conservateur, section de la zoologie, Le Musée du Manitoba, Winnipeg (Manitoba)
Patrick Nantel, Bureau du scientifique en chef des écosystèmes, Parcs Canada, Gatineau (Québec)
Michael Oldham, Centre d'information sur le patrimoine naturel de l'Ontario, ministère des Richesses naturelles de l'Ontario, Peterborough (Ontario)
Suzanne Prange, Furbearer Biologist, Ohio Department of Natural Resources (Ohio)
Brian Rollings, biologiste de la faune, Michigan Department of Natural Resources, Marquette (Michigan)
Rich Russell, Service canadien de la faune, Environnement Canada, Downsview (Ontario)
Michael Schiavone, biologiste de la faune accrédité, New York State Department of Environmental Conservation, Albany (New York)
Irene Schneider, ministère des Richesses naturelles de l'Ontario, Dryden (Ontario)
Sonia Schnobb, adjointe administrative, Secrétariat du COSEPAC, Service canadien de la faune, Environnement Canada, Gatineau (Québec)
Jessica Sicoly, biologiste, gestion des ressources, ministère des Richesses naturelles et des Forêts de l'Ontario, Sault Ste Marie (Ontario)
Don Sutherland, Centre d'information sur le patrimoine naturel de l'Ontario, ministère des Richesses naturelles de l'Ontario, Peterborough (Ontario)
Patrick Tate, Furbearer Project Leader, New Hampshire Fish and Game, Concord (New Hampshire)
John Van den Broeck, biologiste des espèces en péril, biodiversité, ministère des Richesses naturelles de l'Ontario, Fort Frances (Ontario)
Bill Watkins, zoologiste, conservation de la biodiversité, Direction de la faune, Conservation et Gestion des ressources hydriques Manitoba, Winnipeg (Manitoba)
Jenny Wu, chargée de projet scientifique, Secrétariat du COSEPAC, Service canadien de la faune, Environnement Canada, Gatineau (Québec)
Anderson, R.M. 1939. Mammifères de la province de Québec (révisé). The Provencher Society of Natural History of Canada, 1939 Annual Report. pp.37-111.
Anon. 2012. Scent camera station protocol - Pelee Island Grey Fox project, 8 May 2012. Nature Conservancy of Canada.
Arscott, C., comm. pers. 2009. Correspondance par courriel adressée à Environnement Canada (site Web du Registre public des espèces en péril), 1er septembre 2009, Catchacoma, Ontario.
Ashton, K.G., M.C. Tracy et A. deQueroz. 2000. Is Bergmann's rule valid for mammals? American Naturalist 156: 390-415.
Assell, R. 2004. Lake Erie Ice Cover Climatology 1898-2002. NOAA Technical Memorandum GLERL - 128, disponible à l'adresse : PDF. [consulté en août 2015].
Aubry, K. B., M.J. Statham, B.N. Sacks, J.D. Perrine et S.M. Wisely. 2009. Phylogeography of the North American red fox: Vicariance in Pleistocene forest refugia. Molecular Ecology 18: 2668-2686.
Bachmann, comm. pers. 2014. Correspondance par courriel adressée à J. Bowman. Novembre 2014. Senior Wildlife Research Technician, Ontario Ministry of Natural Resources and Forestry, Peterborough, Ontario.
Bachmann, P. et W. Lintack. 1982. Movement and activity of a grey fox (Urocyon cinereoargenteus) in Ontario. Ontario Ministry of Natural Resources Rabies Research Unit Unpublished Report. 5 pp.
Banfield, A.W.F. 1974. The Mammals of Canada. University of Toronto Press, Toronto. xxv + 437 pp. (Également disponible en français : Banfield, A.W.F. 1974. Les mammifères du Canada, Presses de l'Université Laval, Québec, 406 p.)
Bangs, O. 1899. A new Gray Fox from the upper Mississippi valley. Proceedings of the New England Zoological Club 1: 43-33.
Berezanski, D., comm. pers. 2014. Correspondance par courriel adressée à J. Pearce. Février 2014. Provincial Furbearer Biologist, Manitoba Conservation and Water Stewardship, Winnipeg, Manitoba.
Bernier, C., comm. pers. 2014. Conversation téléphonique avec J. Pearce. Juillet 2014. Furbearer Biologist, Vermont Fish and Wildlife, Springfield, Vermont.
Bezdek, H. 1944. A red-grey fox hybrid. Journal of Mammalogy 25:90.
Bowman, J., comm. pers. 2015. Commentaires écrits sur l'ébauche du rapport adressés à Jennie Pearce, avril 2015. Ontario Ministry of Natural Resources, Peterborough, Ontario.
Bowman, J., T.L. Pulfer, L. Bruce, M. McFarlane, C. Sadowski, J. Croswaithe et J. Nageleisen. 2013. Gray Fox (Urocyon cinereoargenteus) monitoring in Pelee Island. Rapport inédit. Ontario Ministry of Natural Resources, Peterborough, Ontario. 2pp.
Bozarth, C.A., S.L. Lance, D.J. Civitello, J.L. Glenn et J.E. Maldonado. 2011. Phylogeography of the gray fox (Urocyon cinereoargenteus) in the eastern United States. Journal of Mammalogy 92: 283-294.
Bruce, L., comm. pers. 2015. Résultats des autopsies de renards gris communiqués à Jennie Pearce. Ministry of Natural Resources and Forestry, Peterborough, Ontario.
Campbell, C.A., A.I. Dagg, M. Dyer et M.E. Gartshore. 1972. Mammals of Waterloo and South Wellington Counties 1972. Impression hors commerce, Waterloo, ON. 130 pp.
Canadian Endangered Species Conservation Council (CESCC). 2006.
Wild Species 2005: The General Status of Species in Canada. (Également disponible en français : Conseil canadien pour la conservation des espèces en péril. 2006. Les espèces sauvages 2005 : Situation générale des espèces au Canada. PDF.)
Canadian Endangered Species Conservation Council (CESCC). 2011. Wild Species 2010: The General Status of Species in Canada. National General Status Working Group: 302 pp. [consulté le 17 juin 2014]. (Également disponible en français : Conseil canadien pour la conservation des espèces en péril. 2011. Espèces sauvages 2010 : La situation générale des espèces au Canada. Groupe de travail national sur la situation générale. 323 p. PDF.)
Chamberlin, M.J. et B.D. Leopold. 2000. Spatial use patterns, seasonal habitat selection, and interactions among adult gray foxes in Mississippi. Journal of Wildlife Management 64: 742-751.
Cormier, J., comm. pers. (2014). Correspondance par courriel adressée à J. Pearce. Novembre 2014. Furbearer Management Biologist, Department of Natural Resources, Fredericton, New Brunswick.
COSEWIC. 2002. COSEWIC assessment and update status report on the grey fox Urocyon cinereoargenteus interior in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. vi + 32 pp. (Également disponible en français : COSEPAC. 2002. Évaluation et Rapport de situation du COSEPAC sur le renard gris (Urocyon cinereoargenteus) au Canada - Mise à jour. Comité sur la situation des espèces en péril au Canada, Ottawa, vi + 34 p.)
Crooks, K.R. et M.E. Soulé. 1999. Mesopredator release and avifaunal extinctions in a fragmented system. Nature 400: 563-566.
Cypher, B.L. 1993. Food item use by three sympatric canids in southern Illinois. Transactions of the Illinois State Academy of Science 86: 139-144.
Cypher, B.L. 2003. Foxes (Vulpes species, Urocyon species, and Alopex lagopus). Pp 511-546. in G.A. Feldhamer, B.C. Thompson et J.A. Chapman (eds.). Wild Mammals of North America: Biology, Management and Conservation. John Hopkins University Press, Baltimore, Maryland.
Cypher, B.L., T.K. Fuller et R. List. 2008. Urocyon cinereoargenteus in IUCN 2013. IUCN Red List of Threatened Species. Version 2013.2. [Anglais seulement, consulté le 26 avril 2014].
Dalpé-Charron, E., comm. pers. 2014. Correspondance par courriel adressée à J. Pearce. Biologiste et coordonnateur provincial, Ministère des Forêts, de la Faune et des Parcs du Québec, Québec, Québec.
Davidson, W.R., V.F. Nettles, L.E. Hayes, E.W. Howerth et C.E. Couvillion. 1992. Diseases diagnosed in gray foxes (Urocyon cinereoargenteus) from the southeastern United States. Journal of Wildlife Diseases 28: 28-33.
Davis, W.B. 1960. The Mammals of Texas (Bulletin 41). Texas Game and Fish Commission, Austin, Texas.
Davis, D.E. et J.E. Wood. 1959. Ecology of foxes and rabies control. Public Health Report 74: 115-118.
DePue, J., comm. pers. 2014. Correspondance par courriel adressée à J. Pearce. Mai 2014. Furbearer/Small Mammal Biologist, Maine Inland Fisheries and Wildlife, Bangor, Maine.
Deschamps, Michael., comm. pers. Septembre 2015. Correspondance par courriel adressée à Graham Forbes. Area Biologist, Ontario Ministry of Natural Resources, Thunder Bay.
Dorf, E. 1959. Climatic changes of the past and present. Contributions from Museum of Paleontology, University of Michigan 8:181-210.
Downing, S.C. 1946. The history of the grey fox in Ontario. Canadian Field-Naturalist 60: 45-46.
Ennen, J., M.E. Kalis, A.L. Patterson, B.R. Kreiser, J.E. Lovich, J. Godwin et C.P. Qualls. 2014. Clinal variation or validation of a subspecies? A case study of the Graptemys nigrinoda complex. Biological Journal of the Linnean Society. 111: 810-822.
Erb, J., comm. pers. 2014. Conversation téléphonique avec J. Pearce. Mai 2014. Furbearer Research Biologist, Minnesota Department of Natural Resources, Grand Rapids, Minnesota.
Essex Region Conservation Authority 2011. Watershed characterization. Updated/Amended Proposed Assessment Report. Essex Region Conservation Authority. PDF. [Anglais seulement, consulté en juillet 2014].
Farias, V., T.K. Fuller, R.K. Wayne et R.M. Sauvajot. 2005. Survival and cause-specific mortality of gray foxes (Urocyon cinereoargenteus) in southern California. Journal of Zoology 266: 249-254.
Fedriani, J. M., T. K. Fuller, R. M. Sauvajot et E. York. 2000. Competition and intraguild predation among three sympatric carnivores. Oecologia 125: 258-270.
Follmann, E.H. 1973. Comparative ecology and behavior of red and gray foxes. Thèse de doctorat, University of Carbondale, Southern Illinois, IL.
Follmann, E.H. 1978. Annual reproductive cycle of the male gray fox. Transactions of the Illinois State Academy of Science 71: 304-311.
Fritzell, E.K. 1987. Gray fox and island gray fox. pp. 409-420 in M. Novak, J.A. Baker, M.E. Obbard et B. Malloch (eds.). Wild Furbearer Management and Conservation in North America. Ontario Trappers Association, North Bay, ON. viii + 1150 pp.
Fritzell, E.K. et K.J. Haroldson. 1982. Urocyon cinereoargenteus. Mammalian Species 189: 1-8.
Fuller, T. 1978. Variable home rang sizes of female Gray Foxes. Journal of Mammalogy 59:447-448.
Gauthier, I., comm. pers. 2014. Correspondance par courriel adressée à G. Forbes. Mars 2015. Biologiste, coordonnatrice provinciale, espèces fauniques menacées et vulnérables, Ministère des Forêts, de la Faune et des Parcs, Québec, Québec.
Gier, H.T. 1948. Rabies in the wild. Journal of Wildlife Management 12: 142-153.
Grinnell, J., J.S. Dixon et J.M. Linsdale. 1937. Fur-bearing Mammals of California, Volume II. University of California Press, Berkeley, CA. xiv + 777 pp.
Hallberg, D.L. et G.R. Trapp. 1984. Gray fox temporal and spatial activity in a riparian-agricultural zone in California's Central Valley. pp. 920-928 in R.W. Warner et K.M. Hendrix (eds.). Proceedings of the California riparian systems conference. University of California Press, Berkeley California.
Hall, E.R. 1981. The Mammals of North America, Volume II. John Wiley & Sons, New York, NY. xv + 1181 + 90 pp.
Hall, E.R. 1984. Geographic variation among brown and grizzly bears (Ursus arctos) in North America. University of Kansas Special Publication 13. Lawrence, Kansas.
Hamilton, W.J., Jr. 1943. The Mammals of Eastern United States. Comstock Publishing Co., Ithaca, NY.
Haroldson, K.J. et E.K. Fritzell. 1984. Homes ranges, activity, and habitat use by gray foxes in an oak-hickory forest. Journal of Wildlife Management 48: 222-227.
Hatfield, D.M. 1939. Winter food habits of foxes in Minnesota. Journal of Mammalogy 20: 202-206.
Henke, S.E. et F.C. Bryant. 1999. Effects of coyote removal on the faunal community in western Texas. Journal of Wildlife Management 63: 1066-1081.
Hockman, G. et J. Chapman. 1983. Comparative feeding habits of red foxes (Vulpes vulpes) and gray foxes (Urocyon cinereoargenteus) in Maryland. American Midland Naturalist 110:276-285.
IUCN 2010. Guidelines for application of IUCN Red List Criteria at regional and national levels. Version 4.0. Prepared by the IUCN Species Survival Commission, January 2010. (Également disponible en français : UICN 2010. Lignes directrices pour l'application des critères de la Liste rouge de l'UICN aux niveaux régional et national. Version 4.0. Préparé par la Commission de la sauvegarde des espèces de l'UICN, janvier 2010.)
IUCN 2014. Guidelines for Using the IUCN Red List Categories and Criteria. Version 11. Prepared by the Standards and Petitions Subcommittee. Disponible à l'adresse : PDF (Anglais seulement).
Jackson, H.H.T. 1961. Mammals of Wisconsin. University of Wisconsin Press, Madison, WI. xii + 504 pp.
James, F. 2010. Avian subspecies: Introduction. Ornithological Monographs 67:1-5.
Jaslow, C.R. 1987. Morphology and digestive efficiency of red foxes (Vulpes vulpes) and grey foxes (Urocyon cinereoargenteus) in relation to diet. Canadian Journal of Zoology 65: 72-79.
Jolicoeur, H., P-Y. Collin, N. Latour et P. Canac-Parquis. 2011. Mise à jour des mentions de renard gris (Urocyon cinereargenteus) dans le sud du Québec. Le Naturaliste Canadien 135: 74-81.
Judge, K. et M. Haviernick. 2002. Update status report on the grey fox Urocyon cinereoargenteus in Canada. COSEWIC 32 pp. (Également disponible en français : Judge, K.A. et M. Haviernick. 2002. Rapport de situation du COSEPAC sur le renard gris (Urocyon cinereoargenteus) au Canada - Mise à jour, in Évaluation et Rapport de situation du COSEPAC sur le renard gris (Urocyon cinereoargenteus) au Canada - Mise à jour. Comité sur la situation des espèces en péril au Canada, Ottawa, 34 p.)
Kutschera, V., N. Lecomte, A. Janke, N. Selva, A.A. Sokolov, T. Haun, K. Steyer, C. Nowak et F. Haller. 2013. A range-wide synthesis and timeline for phylogeographic events in the red fox (Vulpes vulpes). BMC Evolutionary Biology 13:114.
Latham, R.M. 1943. An ecological study of the red and gray foxes in southeastern Pennsylvania (Pittman-Robertson Report I-R). Pennsylvania Game Commission, Harrisburg, Pennsylvania.
Latimer, J. 2014. Northern Minnesota phenology for February 2014: Gray Fox. [Anglais seulement, consulté en juin 2014]
Layne, J.N. 1958. Reproductive characteristics of the gray fox in southern Illinois. Journal of Wildlife Management 22: 157-163.
Layne, J.N. et W.H. McKeon. 1956a. Notes on red and gray fox den sites in New York. New York Fish and Game Journal 3: 248-249.
Layne, J.N. et W.H. McKeon. 1956b. Some aspects of red fox and gray fox reproduction in New York. New York Fish and Game Journal 3: 44-74.
Lindbald-Toh, K. (et 46 coauteurs). 2005. Genome sequence, comparative analysis and haplotype structure of the domestic dog. Nature 438:803-819.
Linnitt, A., comm. pers. 2009. Correspondance par courriel adressée à Environnement Canada (adresse de demande de renseignements généraux). 1er juin 2009, Minto, Ontario.
London Free Press. 1998. Rare grey fox discovered. London Free Press, Monday, 15 June 1998.
Lord, R.D., Jr. 1961. A population study of the gray fox. American Midland Naturalist 66: 87-109.
MacDonald, I.D. 1989. A Biological Inventory and Evaluation of the Oriskany Sandstone and Woodlands Area of Natural and Scientific Interest. Parks and Recreational Areas Section, OMNR, OFER 8903, Central Region, Richmond Hill, ON. x + 193 pp.
Maine Department of Inland Fisheries and Wildlife (MEDIFW) 2013. Maine Hunting and Trapping. Maine Department if Inland Fisheries and Wildlife. [Anglais seulement, consulté le 1er juillet 2014].
Manitoba Wildlife Branch. 2015. Fur bearing animal harvest and values. Wildlife Branch. Government of Manitoba. [Anglais seulement, consulté en août 2015].
McAlpine, D., comm. pers. 2014. Correspondance par courriel adressée à J. Pearce. Juin 2014. Head, Department of Natural Science, Research Curator and Head, Zoology Section, New Brunswick Museum, Saint John, New Brunswick.
McAlpine, D., J.D. Martin et C. Libby. 2008. First occurrence of the grey fox, Urocyon cinereoargenteus, in New Brunswick: a climate-change mediated range expansion? Canadian Field-Naturalist 122: 169-171.
McAlpine, D.F, J. Cormier et I.G. McLean. (2016). Second report of the Grey Fox, Urocyon cinereoargenteus, in New Brunswick and its conservation significance. Canadian Field-Naturalist.
McFarlane, M., données inédites. 2013. Donnée inédites. The Nature Conservancy of Canada, London, Ontario.
McFarlane, M. 2013. What does the fox say? Well that really wasn't the focus of the study. Nature Conservancy of Canada, Toronto, Ontario. [Anglais seulement, consulté le 21 avril 2015].
Merriam, C.H. 1903. Eight new mammals from the United States. Proceedings of the Biological Society of Washington 16: 73-78.
Merriam, C.H. 1918. Review of the Grizzly and Big Brown Bears of North America. North America Fauna 41:1-136.
Michigan Department of Natural Resources (MIDNR). 2013. 2013 Hunting and Trapping Digest. Michigan Department of Natural Resources, Lancing, MI. Disponible à l'adresse : PDF. [consulté le 1er juillet 2014].
Minnesota Department of Natural Resources (MNDNR) 2014. Mammals of Minnesota: Grey Fox. Minnesota Department of Natural Resources, St. Paul, MN. [Anglais seulement, consulté le 15 mai 2014].
Ministères des Forêts, de la Faune et des Parcs du Québec (MFFP). 2014. Données inédites, Ministeres des Forets, de la Faune et des Parcs du Québec, Québec, Québec.
Ministères des Forêts, de la Faune et des Parcs du Québec (MFFP). 2014a. Trapping in Québec, 2014-2016. [consulté le 21 avril 2015]. (Également disponible en français : Ministère des Forêts, de la Faune et des Parcs du Québec (MFFP). 2014a. Piégeage au Québec : Principales règles - Saison 2014-2016.)
Ministères des Forêts, de la Faune et des Parcs du Québec (MFFP). 2014b. Sport Hunting in Québec 2014-2016. Ministère des Ressources naturelles et de la Faune, Québec City, Québec. Site Web : http://mffp.gouv.qc.ca/english/publications/online/wildlife/hunting-regulations/index.asp. [consulté le 21 avril 2015]. (Également disponible en français : Ministère des Forêts, de la Faune et des Parcs du Québec (MFFP). 2014b. Chasse sportive au Québec : Principales règles 1er avril 2014 au 31 mars 2016. Ministère des Ressources naturelles et de la Faune, Québec, Québec.)
Ministères des Forêts, de la Faune et des Parcs du Québec (MFFP). 2014c. Liste de la faune vertébrée du Québec. [consulté le 22 avril 2014].
Mollhagen, T.R., R.W. Wiley et R.L. Packard. 1972. Prey remains in golden eagle nests: Texas and New Mexico. Journal of Wildlife Management 36: 784-792.
Moore, J.E. 1952. The gray fox in Alberta. Journal of Mammalogy 33: 253.
Nantel, P., comm. pers. 2014. Correspondance par courriel adressée à J. Pearce. Février 2014. Bureau du scientifique en chef des écosystèmes, Parcs Canada, Gatineau, Québec.
NatureServe. 2014. NatureServe Explorer: An online encyclopedia of life [application Web]. Version 7.1. NatureServe, Arlington, Virginia. Site Web : http://www.natureserve.org/explorer. [consulté en février 2014].
New Hampshire Fish and Game Department (NHFGD). 2014. 2013 - 2014 New Hampshire Hunting and Trapping Digest. New Hampshire Fish and Game Department, Concord, NH. [Anglais seulement, consulté le 1er juillet 2014].
Niagara Region Conservation Authority. 2013. Updated Assessment Report: Niagara Peninsula Source Protection Area. Niagara Peninsula Conservation Authority, Welland, ON. [consulté en juillet 2014].
Nicholson, W. S. 1982. An ecological study of the gray fox in east-central Alabama. Mémoire de maîtrise ès sciences, Auburn University, Auburn, Alabama. 93pp.
Nicholson, W.S. et E.P. Hill. 1984. Mortality in gray foxes from east-central Alabama. Journal of Wildlife Management 448: 1429-32.
Nicholson, W.S., E.P. Hill et D. Briggs. 1985. Denning, pup-rearing, and dispersal in the gray fox in east-central Alabama. Journal of Wildlife Management 49: 33-37.
Nowak, R.M. 1995. Another look at wolf taxonomy. Pp 375-397 in L. Carbyn, S. Fritts et D. Seip (eds.). Ecology and Conservation of Wolves in a Changing World. Canadian Circumpolar Institute, Alberta. 620 pp.
New York Department of Environment and Conservation (NYSDEC). 2014. Hunting Seasons. New York State Department of Environmental Conservation [Anglais seulement, consulté le 1er juillet 2014].
NOAA 2015. Lake Erie Ice Cover [Anglais seulement, consulté en août 2015].
Ohio Department of Natural Resources (OHDNR). 2014a. Gray Fox - Urocyon cinereoargenteus. Ohio Department of Natural Resources Division of Wildlife. Site Web : http://wildlife.ohiodnr.gov/species-and-habitats/species-guide-index/mammals/gray-fox. [consulté en août 2015].
Ohio Department of Natural Resources (OHDNR). 2014b. 2014 - 2015 Season Dates & Bag Limits. Ohio Department of Natural Resources Division of Wildlife. [Anglais seulement, consulté le 1er juillet 2014].
Ontario Ministry of Natural Resources 2010. Forest Management Guidelines for Conserving Biodiversity at Stand and Site Scales. Queen's Printer, Toronto, Ontario, 211 pp.
Ontario Partners in Flight. 2008a. Ontario Landbird Conservation Plan: Boreal Hardwood Transition, North American Bird Conservation Region 12. Ontario Ministry of Natural Resources, Bird Studies Canada, Environment Canada. PDF. [consulté le 30 juin 2014].
Ontario Partners in Flight. 2008b. Ontario Landbird Conservation Plan: Lower Great Lakes/St. Lawrence Plain, North American Bird Conservation Region 13. Ontario Ministry of Natural Resources, Bird Studies Canada, Environment Canada. PDF. [consulté le 30 juin 2014].
Ordenana, M. (et 11 coauteurs). Effects of urbanization on carnivore species distribution and richness. Journal of Mammalogy 91:1322-1331.
Palmer, R. S. 1956. Gray fox in the Northeast. Maine Field Naturalist 12:62-70.
Parks Canada. 2014. Managed Area Element Status Assessment. Rapport inédit, Agence Parcs Canada, Ottawa, Ontario.
Petersen, L.R., M.A. Martin et C.M. Pils. 1977. Status of gray foxes in Wisconsin, 1975. Wisconsin Department of Natural Resources, Research Report No. 94, Madison, WI. 17 pp.
Peterson, R.L., R.O. Stanfield, E.H. McEwen et A.C. Brooks. 1953. Early records of the red and the gray fox in Ontario. Journal of Mammalogy 34: 126-127.
Pils, C.M. et W.D. Klimstra. 1975. Late fall foods of the gray fox in southern Illinois. Transactions of the Illinois State Academy of Science 68: 255-262.
Prange, S., comm. pers. 2014. Conversation téléphonique avec J. Pearce. 18 novembre 2014. Furbearer Biologist, Ohio Department of Natural Resources, Ohio.
Pratt, Paul, comm. pers. 2015. Correspondance par courriel adressée à Graham Forbes, juillet 2015.
Rausch, R. 1963. Geographic variation in size of North American Brown Bear (Ursus arctos) as indicated by condylobasal length. Canadian Journal of Zoology 41:33-45.
Richmond, N.D. 1952. Fluctuations in gray fox populations in Pennsylvania and their relationship to precipitation. Journal of Wildlife Management 16: 198-208.
Richards, S.W. et R.L. Hines. 1953. Wisconsin fox populations. Wisconsin Conservation Department, Technical Wildlife Bulletin No. 6, Madison, WI. 78 pp.
Rollings, B., comm. pers. 2014. Conversation téléphonique avec J. Pearce. Juillet 2014. Wildlife Biologist, Michigan Department of Natural Resources, Marquette, Michigan.
Root, D.A. 1981. Productivity and mortality of gray foxes and raccoons in southwestern Wisconsin. Mémoire de maîtrise ès sciences, University of Wisconsin, Stevens Point, Wisconsin.
Root, D.A. et N.F. Payne. 1985. Age-specific reproduction of gray foxes in Wisconsin. Journal of Wildlife Management 49: 890-892.
Sadowski, C., comm. pers. 2015. Results of Grey Fox necropsies reported to Jennie Pearce. Ministry of Natural Resources and Forestry, Peterborough, Ontario.
Schiavone, M., comm. pers. 2014. Correspondance par courriel et conversation téléphonique avec J. Pearce. Mai 2014. Certified Wildlife Biologist, New York State Department of Environmental Conservation, Albany, New York.
Schmeltz, L.L. et J.O. Whitaker, Jr. 1977. Use of woodchuck burrows by woodchucks and other mammals. Transactions of the Kentucky Academy of Science 38: 79-82.
Schreber, J. 1775. Die Säugthiere in Abbildungen nach der Natur mit Beschreibungen. 7 volumes; Plate 92 (Vol. 2); Text 361 (Vol. 3).
Sheldon, W.G. 1949. Reproductive behavior of foxes in New York State. Journal of Mammalogy 30: 236-246.
Sheldon, W.G. 1953. Returns on banded red and gray foxes on New York state. Journal of Mammalogy 34, 125.
Sicoly, J., comm. pers. 2014. Conversation téléphonique avec J. Pearce. Septembre 2014. Management Biologist, Ontario Ministry of Natural Resources and Forestry, Sault Ste Marie, Ontario.
Simmons, J.M., W.S. Nicholson, E.P. Hill et D.B. Briggs. 1980. Occurrence of (Dirofilaria immitis) in gray fox (Urocyon cinereoargenteus) in Alabama and Georgia. Journal of Wildlife Diseases 16: 225-228.
Smith, H.C. 1993. Alberta mammals: an atlas and guide. Provincial Museum of Alberta, Edmonton, AB. xii + 239 pp.
Steelman, H.G., S.E. Henke et G.M. Moore. 2000. Bait delivery for oral rabies vaccine to gray foxes. Journal of Wildlife Diseases 36: 744-51.
Stone, W.B., Jr., E. Parks, B.L. Weber et F.J. Parks. 1972. Experimental transfer of sarcoptic mange from red foxes and wild canids to captive wildlife and domestic animals. New York Fish and Game Journal 19: 1-11.
Sullivan, E.G. 1956. Gray fox reproduction, denning, range, and weights in Alabama. Journal of Mammalogy 37: 346-51.
Sullivan, E.G. et A.O. Haugen. 1956. Age determination of foxes by x-ray of forefeet. Journal of Wildlife Management 36: 166-70.
Sutton, R. 1958. A Manitoba record of the Gray Fox. Journal of Mammalogy 39:439-440.
Tate, P., comm. pers. 2014. Conversation téléphonique avec J. Pearce. Juillet 2014. Furbearer Project Leader, New Hampshire Fish and Game, Concord, New Hampshire.
Trapp, G.R. 1978. Comparative behavioural ecology of the ringtail (Bassariscus astutus) and gray fox (Urocyon cineroargenteus) in southwestern Utah. Carnivore 1: 3-32.
Trapp, G.R. et D.L. Hallberg. 1975. Ecology of the grey fox (Urocyon cinereoargenteus): a review. pp. 164-178 in M.W. Fox (ed.). The Wild Canids: their Systematics, Behavioral Ecology and Evolution. Van Nostrand Reinhold Company, New York, NY. xvi + 508 pp.
Tullar, B.F., Jr. et L.T. Berchielli Jr. 1982. Comparison of red foxes and gray foxes in central New York with respect to certain features of behavior, movement and mortality. New York Fish and Game Journal 29: 127-133.
Van den Broeck, J. 2014a. Records of confirmed Grey Fox (Urocyon cineroargenteus) Northwestern Ontario. Rapport inédit du Ontario Ministry of Natural Resources, Fort Frances, Ontario
Van den Broeck, J. 2014b. Species occurrence record - MNR Fort Frances District, Grey Fox. Rapport inédit du Ontario Ministry of Natural Resources, Fort Frances, Ontario.
Vermont Department of Fish and Wildlife (VTDFW). 2014. 2014 Vermont Hunting, Fishing and Trapping Laws and Guide: Fish and Wildlife Regulations. Vermont Department of Fish and Wildlife. Site Web : http://www.eregulations.com/vermont/pageFlip/. [consulté le 1er juillet 20114].
Waters, J.H. 1964. Red fox and gray fox from New England archeological sites. Journal of Mammalogy 45: 307-308.
Waters, J.H. 1967. Foxes on Martha's Vineyard, Massachusetts. Journal of Mammalogy 48: 137-138.
Weston Glenn, J., D. Civitello et S. Lance. 2009. Multiple paternity and kinship in the gray fox (Urocyon cinereoargenteus). Mammalian Biology 79:394-402.
Weston, J. et L. Brisbin 2003. Demographics of a protected population of gray foxes (Urocyon cinereoargenteus) in South Carolina. Journal of Mammalogy 84:996-1005.
Wilson, D. E. et D. M. Reeder (editors). 2005. Mammal Species of the World: a Taxonomic and Geographic Reference. Third Edition. John Hopkins University Press, Baltimore, MD. xviii + 2142 pp. [Anglais seulement, consulté en juin 2014].
Wintemberg, W.J. 1921. Archaeological evidence concerning the presence of the grey fox (Urocyon sp.) in Ontario. Canadian Field-Naturalist 35: 19-20.
Wintemberg, W.J. 1928. Uren prehistoric village site, Oxford County, Ontario. National Museum of Canada Bulletin No. 51, Anthropological Series No. 10. 97 pp.
Wintemberg, W.J. 1936. Roebuck prehistoric village site, Grenville County, Ontario. National Museum of Canada Bulletin No. 83, Anthropological Series No. 19. 178 pp.
Wintemberg, W.J. 1939. Lawson prehistoric village site, Middlesex County, Ontario. National Museum of Canada Bulletin No. 94, Anthropological Series No. 25. 104 pp.
Wintemberg, W.J. 1948. The Middleport prehistoric village site, Brant County, Ontario. National Museum of Canada Bulletin No. 109, Anthropological Series No. 27. v+ 79 pp.
Wisconsin Department of Natural Resources (WIDNR). 2013. Small game hunting regulations, 2013 fall turkey, and 2014 spring turkey regulations. Site Web : PDF. [consulté le 1er juillet 2014].
Wong, J. (et 4 coauteurs). Great Lakes Ice Climatology Update: Winter 2006-2011. GLERL-155. PDF. [consulté en août 2015].
Wood, J.E. 1958. Age structure and productivity of a gray fox population. Journal of Mammalogy 39: 74-86.
Wood, J.E., D.E. Davis et E.V. Komarek. 1958. The distribution of fox populations in relation to vegetation in southern Georgia. Ecology 39: 160-162.
Wooding, J.B. 1984. Coyote food habits and the spatial relationship of coyotes and foxes in Mississippi and Alabama. Mémoire de maîtrise ès sciences, Mississippi State University, State College.
Wooding, S. et R. Ward. 1997. Phylogeography and Pleistocene evolution in the North American black bear. Molecular Biology and Evolution 14:1096-1105.
Yearsley, E.F. et D.E. Samuel. 1980. Use of reclaimed surface mines by foxes in West Virginia. Journal of Wildlife Management 44: 729-734.
Yuan Chung Zee. 1999. Rhabdoviridae. pp. 412-417 in D.C. Hirsh et Yuan Chung Zee (eds.). Veterinary Microbiology. Blackwell Science Inc., Malden, MA. vii + 479pp.
Mme Jennie L. Pearce (Ph. D.) est née en Australie et a émigré au Canada en 1999. Dans ces deux pays, ses recherches ont mis l'accent sur la modélisation des exigences liées à la répartition, à la viabilité et à l'habitat des espèces sauvages pour documenter et orienter les mesures de conservation. Mme Pearce a mis au point des modèles de répartition spatiale visant des espèces de mammifères dont les mentions de répartition sont limitées, a développé des modèles de viabilité des populations pour des espèces à petits effectifs et a modélisé les changements démographiques temporels susceptibles de se produire dans les cas où les paysages sont modifiés par les activités humaines. Elle a rédigé ou participé à la rédaction de 10 rapports du COSEPAC et publié plus de 40 articles scientifiques dans le domaine de la biologie de la conservation.
Aucune collection n'a été examinée.
| Date | Localité | Observateur | Source |
|---|---|---|---|
| Date | Localité | Observateur | Source |
| 1946-1947 | Sprague | A. Suchrov | Sutton (1958) |
| 12 janvier 1957 | St. Adolphe | M. Bouchard | Dossiers du Musée Manitobain de l'homme et de la nature; Sutton (1958) |
| 9 décembre 1977 | À 7 milles au sud et à 2 milles à l'ouest de Hadishville | E. Shindler | Dossiers du Musée Manitobain de l'homme et de la nature |
| 20 octobre 2000 | À 5 milles au nord de Piney | P. Krahn | Dossiers du Musée Manitobain de l'homme et de la nature |
| Vers 2002 | Whiteshell (individu piégé) | - | Berezanski (comm. pers.) |
| Tous les deux ans | Région de Sprague (mortalité routière) | - | Berezanski (comm. pers.) |
| Individus | Date | Localité | Source (observateur) | Numéro de l'observation au CIPN |
|---|---|---|---|---|
| 1-2 | 1er janvier 1944 2 octobre 1944 |
Comté de Rainy River, lac Wild Potato (ROM16311) | Dossiers du MRO (S. Boy) | 219386, 219387 |
| 1-2 | 22 février 1982 | Comté de Thunder Bay, juste au nord et à l'ouest du lac Shebandowan | M. Lankester, comm. pers. (V. Hollet) | 219385, 219565 |
| 1-2 | 10 décembre 1991 | Comté de Thunder Bay, au sud du lac Dog | B. Wall, comm. pers. (B. Wall) | 219384, 219383 |
| 1 | Décembre 1997 | Comté de Rainy River, à 5 km au sud-est d'Atikokan | Individu piégé, B. Jackson, comm. pers. (un piégeur) | 219564 |
| 1 mâle | Février 2006 | Au nord-ouest de Fort Frances, à 800 m au sud du lac Little Pine | Individu piégé, Van den Broeck (2014a) | - |
| 1 mâle | 2008a | Inlet Rocky | Individu piégé, Van den Broeck (2014a) | - |
| 1 mâle | 2008b | Couchiching | Individu piégé, Van den Broeck (2014a) | - |
| 1 | Novembre 2010 | Chemin du lac Mountain, Thunder Bay | Photo, Van den Broeck (2014a) | - |
| 3 | 11 février 2011 | Chemin Fern, lac Whitefish, Thunder Bay | Photo, Van den Broeck (2014a) | - |
| 1 | 9 février 2011 | Lac Calm, Atikokan | Photo, Van den Broeck (2014a) | - |
| 1 | 5-7 mai 2012 | Péninsule Sibley, Observatoire des oiseaux du cap Thunder | Individu observé, Van den Broeck (2014a) | - |
| 1 | 2012 | Mine centre, district de Fort Francis | Individu piégé, Van den Broeck (2014a) | - |
| 2 | Hiver 2012 | Chemin Mountain, Thunder Bay | Photo, Van den Broeck (2014a) | - |
| 4 | Été/automne 2013 | Chemin Mountain, Thunder Bay | Individu observé, puis un individu tué sur la route, Van den Broeck (2014a) | - |
| 1 mâle | Novembre 2013 | Chemin Delints - près de la région du lac White Fish | Individu piégé, Van den Broeck (2014a) | - |
| 1 mâle | Hiver 2014 | Dorion | Individu observé et mort par la suite, Van den Broeck (2014a) | - |
| 1 mâle, 1 femelle | 2014 | Rainy River, district de Fort Francis | Mortalité routière, Van den Broeck (2014a) | - |
| 1 mâle | 2014 | Thunder Bay, chemin Devon - près de la frontière | Individu piégé, Van den Broeck (2014a) | - |
| 1 femelle en lactation | Été 2015 | Chemin Mountain, Thunder Bay | B. Ratcliff (photo) | - |
| Individus | Date | Localité | Source (observateur) | Numéro de l'observation au CIPN |
|---|---|---|---|---|
| 1-4 | Octobre 1983 | Comté d'Essex, île Pelée à l'altitude maximale | Harris (1994) (L. Defraeve) | 79378 |
| 1 | Octobre 1983 | Comté d'Essex, île Pelée à la pointe nord | Harris (1994) (L. Defraeve) | 79379 |
| 1 | 1er novembre 1984 | Comté d'Essex, île Pelée, alvar du chemin Stone (ROM91445) | Dossiers du MRO (T. Gardner) | 79386 |
| 1 | Avril 1985 | Comté d'Essex, île Pelée, individu mort sur la route | D. Coulson, comm. pers. (D. Coulson, C. Campbell) | 79377 |
| 1 | 31 janvier 1989 | Comté d'Essex, île Pelée, moitié sud | Harris (1994) (K. Hillman) | 79385 |
| 1 | Vers 1990 | Comté d'Essex, île Pelée | N. Beattie, comm. pers. (N. Beattie) | 79376 |
| 1 | 1994 | Comté d'Essex, île Pelée, au nord de la carrière | B. Porchuk, comm. pers. (B. Porchuk) | 79375 |
| 2 | Novembre 1996 | Comté d'Essex, île Pelée, sur le chemin dans la réserve naturelle de la pointe Fish (2 individus) | B. Porchuk, comm. pers. (B. Porchuk) | 19374 |
| 1 | Printemps 1997 | Comté d'Essex, île Pelée, sur le terrain de B. Porchuk sur le chemin Curry Dyke, à l'extrémité sud de l'île | B. Porchuk, comm. pers. (B. Porchuk) | 79384 |
| 1 | 1er juin 1998 | Comté d'Essex, île Pelée | London Free Press (1998) (D. Hodare) | 79383 |
| 1 | 7 juin 1998 | Comté d'Essex, île Pelée, sur le chemin dans la réserve naturelle de la pointe Fish | P. Pratt, comm. pers. (P. Pratt) | 79382 |
| 1 | 12 avril 2000 | Comté d'Essex, île Pelée, réserve naturelle de la pointe Fish | B. Porchuk, comm. pers. (B. Porchuk) | 79381 |
| 1 | 11 juin 2000 | Comté d'Essex, île Pelée, à la croisée du chemin Shore Ouest, au nord du quai du traversier | P. Pratt, comm. pers. (Essex Co. Field Naturalists Club) | 79380 |
| 1 | Janvier 2001 | Comté d'Essex, île Pelée | N. Beattie, comm. pers. (N. Beattie) | 79373 |
| 1 | 17 juin 2010 | Chalet de CNC, île Pelée, forêt de la pointe Middle (CNC) | Sam Brinker (photographe) | 219350 |
| 1-16 | Hiver 2012-2013 | 8 localités à l'île Pelée | Bowman et al. (2013) | - |
| 1 | Décembre 2012 | Individu tué sur la route, île Pelée | C. Sadowski, comm. pers. | - |
| Individus | Date | Localité | Source (observateur) | Numéro de l'observation au CIPN |
|---|---|---|---|---|
| 1 | Avant 1900 | Marais Hillman | - | 219549 |
| 1 femelle | 20 février 1948 | Port Colborne | - | 219391 |
| 1 mâle | 26 mars 1949 | Comté de Niagara, St. Catharines, Canton géographique de Grantham (ROM19592) [au nord-ouest des chutes du Niagara] | Dossiers du MRO (B. Demars) | 219396 |
| 1 femelle | 1er janvier 1952 | Comté de Niagara, Canton de Wainfleet (près de Chamber's Corners) (ROM21489) | Dossiers du MRO (A.H. Muma) | 219390 |
| 1 mâle | 2 janvier 1952 | Comté de Niagara, Canton de Fort Érié (ROM21491) | Dossiers du MRO (ministère des Terres et des Forêts de l'Ontario) | 219393 |
| 1 femelle | 15 novembre 1952 | Comté de Niagara, Port Colborne, canton géographique de Humberstone (ROM22501) | Dossiers du MRO (A.H. Muma) | 219392 |
| 1 femelle | 1er janvier 1954 | Comté de Niagara, canton de Niagara Falls, près de Chippawa (ROM25502) | Dossiers du MRO (ministère des Terres et des Forêts de l'Ontario) | 219395 |
| 1 | Juillet 1964 | Comté de Niagara, canton de Grimsby, près du chemin Spring Creek et du chemin Park, non loin de la décharge | G. Meyers, comm. pers. | 219397 |
| 1 | 1965 | Comté de Haldimand-Norfolk, marais Grand River, au sud-est de Dunnville | Gartshore (1987) (W. Houser) | 219388 |
| 1 | Janvier 1978 | Comté de Simcoe, près de Stayner | Dobbyn (1994) (piégeur de la région) | 219351 |
| 1 mâle | 11 octobre 1978 |
Lot no 15, concession no 2, canton de Goderich, comté de Huron | Spécimen no 8344, données de surveillance de la rage du MRNO | - |
| 1 | 25 octobre 1978 | Comté de Haldimand-Norfolk, canton de Norfolk, à 6 km au nord de Cultus, concession no VIII, Walsingham (ROM82842) | Dossiers du MRO (O. Wilson) | 219545 |
| 1-2 | Saison de piégeage 1978-1979 | Comté de Niagara, nord ou Allanburg | Harris (1994) (J. Caldwell) | 219569 |
| 1 | 21 mars 1979 | À 2 km au sud-ouest de Wiarton | - | 219563 |
| 1 femelle | 10 novembre 1979 | Lot no 16, concession no 3, canton de Nottawasaga, comté de Lambton | Données de surveillance de la rage du MRNO | - |
| 1 | Saison de piégeage 1979-1980 | Comté de Niagara, canton de Thorold, individu piégé dans le secteur qu'on appelle maintenant le parc industriel Brock | Harris (1994) (L. Schmaltz) | 219568 |
| 1 | 1980 (et vu depuis 1975-1980) |
Comté de Haldimand-Norfolk, canton de Haldimand, Oriskany Sandstone et Woodlands, à 7 km à l'ouest de Cayuga, environ à 1,5 km au sud de Clanbrassil | MacDonald (1989) (S. Banticks) | 219389 |
| 1 mâle | 15 octobre 1980 | Comté de Lambton, canton d'Enniskillen, à environ 4 km à l'est de Petrolia, lot no 26, concession no 12 | Bachmann et Lintack (1982) (W. Lintack) | 219550 |
| 1 mâle | 1er décembre 1980 | Lot no 34, concession no 8, canton de Lindsay, comté de Bruce | Spécimen no 13577, données de surveillance de la rage du MRNO | - |
| 1 | 15 octobre 1980 | Comté d'Essex, à environ 0,5 à 1 mille au nord ou Cottam et à 4 milles à l'est d'Essex | Harris (1994) (P. Jacques) | 219552 |
| 1 | 1980 1985 | Comté d'Essex, boul. Dominion à Windsor | P. Pratt, comm. pers. (Gary's Taxidermy) | - |
| 1 mâle | 1er avril 1982 | Lot no 29, concession no 13, Enniskillen, comté de Lambton | Spécimen no 15788, données de surveillance de la rage du MRNO | - |
| 1 | 1982 | Comté de Haldimand-Norfolk, canton de Delhi, au nord de Waterford Ponds sur Concession Rd. VI | Gartshore (1987) (R. Bergman) | 219548 |
| 2 | 23 mai 1984 | Comté d'Essex, Windsor, du côté nord de Todd Lane, aux limites de LaSalle et de Windsor (2 adultes) | P. Pratt, comm. pers. (S. Wardle, P. Pratt) | 219381 |
| 1-2 | 4 juin 1984 | À la lisière du boisé, derrière une maison de banlieue, près de la prairie Ojibway, à Windsor | P. Pratt (photo) | 219353 |
| 1 | 1985 | Comté d'Essex, Windsor, limite sud de la zone d'intérêt naturel et scientifique (ZINS) Spring Garden | P. Pratt, comm. pers. (S. Wardle) | 219382 |
| 1 | 27 octobre 1985 | Comté d'Essex, canton d'Anderton, Amherstburg, lot no 42, concession no 1 | Harris (1994) (E. Meloche) | 219553 |
| 1 | 10 juin 1986 | Medonte (Coldwater), comté de Simcoe | Spécimen no 42138, soumission de l'ACIA no 86RABN05294 | - |
| 1 | 29 juillet 1986 | Waterford, comté de Norfolk | Spécimen no 22074, soumission de l'ACIA no 86RABN05294 | - |
| 1 | Novembre 1986? | Comté d'Essex, canton d'Anderton, Amherstburg, lot no 40, concession no 1 | Harris (1994) (E. Meloche) | 219556 |
| 1 | Novembre 1987? | Comté d'Essex, canton d'Anderton, Amherstburg, lot no 40, concession no 1 | Harris (1994) (E. Meloche) | 219557 |
| 1 | Novembre- décembre 1987 |
Comté d'Essex, canton d'Anderton, Amherstburg, lot no 42, concession no 1 | Harris (1994) (E. Meloche)) | 219558 |
| 1 | Novembre- décembre 1987 |
Comté d'Essex, canton d'Anderton, Amherstburg, lot no 42, concession no 1 | Harris (1994) (E. Meloche) | 219559 |
| 1-2 | Décembre 1989 ou janvier 1990 | Comté de Lambton, canton de Sombra, entre la concession no 7 et la concession no 8 | Harris (1994) (G. English) | - |
| 1 | Avant 19906 | Comté d'Essex, canton de Mersea, marais Hillman, spécimen naturalisé observé | P. Pratt, comm. pers. (P. Pratt) | - |
| 1 | 14 novembre 19906 | Comté de Simcoe, canton de Mara, lot no 26, concession no 11 | Harris (1994) (B. Edinger) | - |
| 1 | 9 mai 1991 | Comté de Niagara, 7, Bedford Park, Grimsby | G. Meyers, comm. pers. (G. Meyers) | 219398 |
| 1 | 7 février 1997 | Comté de Wellington, canton de Nichol, à environ 3 km au nord d'Elora dans un boisé près du ruisseau Irvine | D. Coulson, comm. pers. (D. Coulson) | 219561 |
| 1 femelle | 28 octobre 2001 | Lot no 13, concession no 2, Goderich, comté de Huron | Spécimen no 74309, données de surveillance de la rage du MRNO | - |
| 1 | 19 août 2002 | Chemin Cameron Sud, à l'ouest de la rue Dougall, à Windsor | - | En attente |
| 1 | Inconnue | À 1,5 km au sud-est de Craighurst, carcasse ramassée | - | 219405 |
| Individus | Date | Localité | Source (observateur) | Numéro de l'observation au CIPN |
|---|---|---|---|---|
| 1 | Janvier 1942 | Stormont, comtés de Dundas et de Glengarry, canton de Kenyon, à 4 milles à l'ouest d'Alexandria (ROM15707) | - | 219406 |
| 1 | Février 1944 | Comté de Frontenac, canton de Kenebec, à 6 milles à l'est de Kaladar (ROM16214) | Dossiers du MRO (R. Knight) | 219401 |
| 1 femelle | 20 février 1948 | Comté de Niagara, Port Colborne, canton géographique de Humberstone (ROM19494) | Dossiers du MRO (W. Lee) | 219391 |
| 1 femelle | 27 avril 1948 | Comtés de Leeds et Grenville, canton d'Escott (ROM18801) | Dossiers du MRO (B. Reed) | 219570 |
| 1 | 1er juillet 1951 | Comtés de Leeds et Grenville, canton d'Elizabethtown, près de Brockville (ROM21579) | Dossier du MRO (inconnu) | 219399 |
| 1 mâle | 27 novembre 1952 | Comté de Lennox et Addington, canton d'Abinger (ROM22493) | Dossiers du MRO (I. Brown) | 219571 |
| 1 | 1963 | Comté de Wellington, sur la route entre Breslau et Guelph | Campbell et al. (1972) (inconnu) | 219562 |
| 1 femelle | 19 mars 1977 | Comté de Peterborough, à 3 km au nord-est de Cavan (ROM80081) | Dossier du MRO (RD McRae) | 219380 |
| 1 | Janvier 1978 | Comté de Lennox et Addington, sur l'île Wolfe près de la pointe Alexander, concession no 6 | Harris (1994) (R. Fawcett) | 219567 |
| 1 | 31 janvier 1979 | Comté de Lennox et Addington, pointe Holiday, île Wolfe | Harris (1994) (R. Hodge) | 219566 |
| 1 mâle | 24 octobre 1979 | Comté de Durham, canton de Pickering, près de Balsam, à 5 km à l'est de Claremont (ROM82841) | Dossiers du MRO (P. Hall) | 219394 |
| 1 | 31 mai 1979 | Dans le nord de l'île Tar, à environ 2 milles en aval du pont des Mille-îles | - | 219400 |
| 1 | Novembre 1980 | Comté de Renfrew, canton de Lyell | Harris (1994) (B. Coulas) | - |
| 1 | Janvier 1981 | Comté de Lanark, canton de Pakenham, concession no 12 | Harris (1994) (C. Coe) | - |
| 1 | Entre 1983 et 1985 | Comté de Northumberland, canton de Cramahe | Harris (1994) (B. Stratton) | - |
| 1 mâle | 15 décembre 2000 | Comté de Lanark, canton de Drummond, le long de la route 7, près de Perth | Spécimen no 68215, données de surveillance de la rage du MRNO | 219402 |
| 1 | Octobre 2008 | Promenade des îles du Saint-Laurent | Mortalité routière, MRNF (L. Bruce) | - |
| 1 | 5 mars 2014 | À la croisée de la route 7, concession no 9, comté de Lanark | Individu observé, MRNF (S. Thompson) | - |
| 1 | Novembre-décembre 2014 | Rue Mills à Bishop Mills | Mortalité routière, MRNF (R. Ciraco) | - |
| Individus | Date | Localité | Source (validée par) |
|---|---|---|---|
| 1 | 1975 | Saint-Bernard-de-Lacolle | Individu piégé, M. Richard (H. Jolicoeur); S. Thérien, K. Racey |
| 1 mâle | Février 1981 | Saint-Bernard-de-Lacolle | Individu piégé, P. Martin (H. Jolicoeur) |
| 1 femelle | Hiver 1984-1985 | Saint-Bernard-de-Lacolle | Individu piégé, P. Martin (H. Jolicoeur) |
| 1 mâle | 1990-2000 | Saint-Anicet | Individu piégé, A. Lebrun/P. Martin (H. Jolicoeur) |
| 1 femelle | 1990-2000 | Saint-Anicet | Individu piégé, A. Lebrun/P. Martin (H. Jolicoeur) |
| 1 | 23 octobre 2003 | Saint-Stanislas-de-Kostka | Individu piégé, A. Lebrun/P. Martin (H. Jolicoeur) |
| 1 mâle | 15 décembre 2007 | Saint-Rémi-de-Napierreville | Individu piégé, D. Tremblay/P. Martin (H. Jolicoeur) |
| 1 | 2009 | Hemmingford | Individu observé, S. Thérien/P. Martin (H. Jolicoeur) |
| 1 | 2009 | Hemmingford | Individu observé, S. Thérien/P. Martin (H. Jolicoeur) |
| 1 | 8 novembre 2010 | Saint-Bernard-de-Lacolle | Individu piégé vivant, P. Martin (H. Jolicoeur) |
| 1 femelle | 2012 | Saint-Bernard-de-Lacolle | Mortalité routière, P. Martin (G. Tremblay) |
| 1 mâle | 30 novembre 2012 | Saint-Bernard-de-Lacolle | Individu piégé, P. Martin (G. Tremblay) |
| 1 mâle | 2013 | Hemmingford | Individu piégé, P. Martin (G. Tremblay) |
| 1 | 12 janvier 2012 | Léry | Individu piégé, S. Lessard [APF] (E. Dalpé-Charron/F. Landry) |
| 1 | 2012 | Saint-Bernard-de-Lacolle | Individu piégé, C. Lemay [APF] (E. Dalpé-Charron/F. Landry) |
| 1 | 28 novembre 2013 | Hemmingford | Individu piégé, P. Martin (E. Dalpé-Charron/F. Landry) |
| 1 | 5 janvier 2014 | Hemmingford | Individu piégé, S. Richard (E. Dalpé-Charron/F. Landry) |
| 1 | 11 février 2014 | Hinchinbrooke | Individu piégé, A. Lebrun (E. Dalpé-Charron/F. Landry) |
| 1 | 2014 | La Prairie | Vidéo (P. Blais) |
| 1 | 2014 | La Prairie | Vidéo (P. Blais) |
| Individus | Date | Localité | Source (validée par) |
|---|---|---|---|
| 1 | 1894 | Cookshire-Eaton (Johnville) | Individu observé, K Racey (H. Jolicoeur) |
| 1 femelle juvénile | 12 décembre 2011 | Sherbrooke | Mortalité routière, M-C Benoît (G. Tremblay) |
| 1 | 8 février 2012 | Cookshire-Eaton | Individu piégé, M. Rolfe (E. Dalpé-Charron/F. Landry) |
| 1 | 1er janvier 2014 | Saint-Malo | Individu piégé, M. Blais (E. Dalpé-Charron/F. Landry) |
| 1 | 5 décembre 2013 | Sherbrooke | Individu piégé, M. Grondin (E. Dalpé-Charron/F. Landry) |
| Individus | Date | Localité | Source (validée par) |
|---|---|---|---|
| 1 | 1940-1950 | Saint-Augustin-de-Woburn | Individu piégé (H. Jolicoeur) |
| 1 | 19 décembre 2011 | Lac-Mégantic | Mortalité routière?, A. Gauthier [APF] (E. Dalpé-Charron/F. Landry) |
| 1 | 12 février 2013 | Frontenac | Individu piégé, Y. Poirier (E. Dalpé-Charron/F. Landry) |
| 1 | 14 novembre 2013 | Saint-Augustin-de-Woburn | Individu piégé, A. Bédard (E. Dalpé-Charron/F. Landry) |
| Individus | Date | Localité | Source (validée par) |
|---|---|---|---|
| 1 | 1er décembre 1986 | Saint-Benjamin | Individu piégé, G et F Caron (H. Jolicoeur) |
| 1 mâle adulte | Automne 1996 | Sainte-Justine | Individu piégé, E. Houle (H. Jolicoeur) |
| 1 mâle adulte | 15 novembre 2005 | Sainte-Aurélie | Individu piégé, J-F Caouette/G. Fortin (H. Jolicoeur) |
| 1 | Décembre 2006 | Saint-Côme-Linière | Individu piégé, G. Poulin (H. Jolicoeur) |
| 1 mâle adulte | 2007 | Saint-Benoît-Labre | Individu piégé, B. Pouliot (H. Jolicoeur) |
| 1 mâle adulte | 22 décembre 2014 | Voir Griffe | Individu piégé, E. Dalpé-Charron |
| 1 mâle | 20 novembre 2014 | Montmagny | Individu piégé, E. Dalpé-Charron |
| Individus | Date | Location | Source |
|---|---|---|---|
| 1 mâle juvénile | 29 octobre 2007 | Rollingdam, comté de Charlotte | Individu piégé (McAlpine et al., 2008) |
| 1 | 19 septembre 2014 | Lac Oromocto, comté de York | Appareil photo actionné par le mouvement, (McAlpine et al., présenté) |
| Impact des menaces | Impact des menaces (descriptions) | Comptes des menaces de niveau 1 selon l'intensité de leur impact : Maximum de la plage d'intensité |
Comptes des menaces de niveau 1 selon l'intensité de leur impact : Minimum de la plage d'intensité |
|---|---|---|---|
| A | Très élevé | 0 | 0 |
| B | Élevé | 1 | 1 |
| C | Moyen | 0 | 0 |
| D | Faible | 0 | 5 |
| - | Impact global des menaces calculé : | Élevée | Élevée |
| Numéro de la menace | Menace | Impact (calculé) |
Impact (calculé) Description |
Portée (10 prochaines années) |
Gravité (10 ans ou 3 générations) |
Immédiateté | Commentaires |
|---|---|---|---|---|---|---|---|
| 1 | Développement résidentiel et commercial | - | - | - | - | - | Aucun signe de menace |
| 2 | Agriculture et aquaculture | - | - | - | - | - | Aucun signe de menace. Facteur limitatif possible (même chose pour l'agriculture et peut-être les incendies) |
| 3 | Production d'énergie et exploitation minière | - | - | - | - | - | Aucun signe de menace |
| 4 | Corridors de transport et de service | - | Ne constituent pas une menace | Généralisée (71-100 %) |
Neutre ou avantage possible | Élevée (continue) |
- |
| 4.1 | Routes et voies ferrées | - | Ne constituent pas une menace | Généralisée (71-100 %) |
Neutre ou avantage possible | Élevée (continue) |
* Peu probable, mais menace inconnue si la mortalité routière est responsable du déclin de la population * Dans la zone d'occurrence, la mortalité routière limite probablement l'expansion de l'espèce; si l'on considère seulement la population reproductrice, l'île Pelée constitue la localité où la population est la plus touchée (> 30 %) Certains membres de l'équipe des évaluateurs des menaces croient que la gravité de la menace dégrade ou réduit les occurrences touchées plutôt que d'être neutre ou d'entraîner un avantage possible |
| 5 | Utilisation des ressources biologiques | B | Élevé | Généralisée - grande (31-100 %) |
Élevée (31-70 %) |
Élevée (continue) |
- |
| 5.1 | Chasse et capture d'animaux terrestres | B | Élevé | Généralisée - grande (31-100 %) |
Élevée (31-70 %) |
Élevée (continue) |
Quel pourcentage des individus sont exposés? Ils sont tous exposés. * il y a eu des prises accessoires aussi loin au nord que North Bay * île Pelée : il y a eu un plus grand nombre d'individus tués sur la route que d'individus piégés * Québec : l'espèce fait l'objet d'un piégeage assez important * en raison de l'incertitude, la portée est de « généralisée à grande » L'espèce peut facilement prendre de l'expansion si elle ne fait pas l'objet de piégeage * aux États-Unis, l'espèce a un taux de mortalité élevé (en raison de la récolte, de la mortalité routière), mais la population maintient un taux de fécondité élevé |
| 5.3 | Exploitation forestière et récolte du bois | - | - | - | - | - | Facteur limitatif possible (même chose pour l'agriculture et peut-être les incendies) * l'espèce n'apprécie pas les milieux très ouverts |
| 6 | Intrusions et perturbations humaines | - | - | - | - | - | Aucun signe de menace |
| 7 | Modifications des systèmes naturels | - | - | - | - | - | - |
| 7.1 | Incendies et suppression des incendies | - | - | - | - | - | Facteur limitatif possible (même chose pour l'agriculture et peut-être les incendies) |
| 8 | Espèces et gènes envahissants ou autrement problématiques | - | Inconnu | Généralisée (71-100 %) |
Inconnue | Modérée (peut-être à court terme, < 10 ans) |
- |
| 8.1 | Espèces exotiques (non indigènes) envahissantes | - | Inconnu | Généralisée (71-100 %) |
Inconnue | Modérée (peut-être à court terme, < 10 ans) |
La maladie de Carré est considérée comme non indigène * quel pourcentage des individus seraient exposés? Ils seraient tous exposés. * est-ce qu'on nous a déjà remis un renard gris qui était mort à la suite de la maladie de Carré? non |
| 8.2 | Espèces indigènes problématiques | - | Inconnu | Grande (31-70 %) |
Inconnue | Élevée (continue) |
La rage est considérée comme indigène * L'immédiateté est « élevée » en raison de 2 cas de mortalité signalés en Ontario |
| 9 | Pollution | - | - | - | - | - | Aucun signe de menace |
| 10 | Phénomènes géologiques | - | - | - | - | - | Aucun signe de menace |
| 11 | Changements climatiques et phénomènes météorologiques violents | - | - | - | - | - | Le réchauffement pourrait être profitable pour l'espèce, d'après la récente expansion de l'aire de répartition vers le nord aux États-Unis |
Détails de la page
- Date de modification :