
COSEWIC Assessment and Status Report on the Gray Fox Urocyon cinereoargenteus in Canada - 2015

Gray Fox - Photo: John Woodcock.
- Document Information
- COSEWIC Assessment Summary
- COSEWIC Executive Summary
- Technical Summary
- Preface
- COSEWIC History
- COSEWIC Mandate
- COSEWIC Membership
- Definitions (2015)
- Wildlife Species Description and Significance
- Distribution
- Habitat
- Biology
- Population Sizes and Trends
- Threats and Limiting Factors
- Protection, Status and Ranks
- Acknowledgements and Authorities Contacted
- Information Sources
- Biographical Summary of Report Writer
- Collections Examined
- Figure 1. A Gray Fox, Urocyon cinereoargenteus, photographed May 2012 at Thunder Cape, northwestern Ontario.
- Figure 2. Range of Gray Fox (Urocyon cinereoargenteus) in mainland North America indicating locations of 8 of 15 subspecies. Gray Fox occur from Canada to Colombia with Canadian breeding sub-populations in the Rainy River - Thunder Bay ('Northwestern Ontario') and Pelee Island areas of Ontario, and non-breeding sub-populations in Manitoba and Quebec. Dispersing individuals have been recorded in Alberta and New Brunswick (not shown). Subspecies shown are: 1) borealis; 2) cinereoargenteus; 3) floridanus; 4) ocythous; 5) scotti; 6) catalinae; 7) californicus; and 8) peninsularis (Hall 1981). Genetic analyses suggest that subspecific designations likely do not exist in eastern North America (Bozarth et al. 2011); in this status report, a single designatable unit is recognized for Gray Fox found in Canada.
- Figure 3. Extent of occurrence (EOO) of Gray Fox (Urocyon cinereoargenteus), based on recent (i.e., 1993 - 2014) records (indicated by stars) of Gray Fox in Manitoba, Ontario, Quebec and New Brunswick. Historical records (< 1993) indicated by triangles. (Map by Jenny Wu.)
- Appendix 1. Details on Observations of Gray Fox by Region, Canada.
- Table A1. Gray Fox records in Manitoba. Source: Judge and Haviernick (2002), D. Berezanski (unpub. data.)
- Table A2. Gray Fox records in Northwestern Ontario. Source: Judge and Haviernick (2002), NHIC data, and Van den Broeck (2014a). Unconfirmed reports are not included.
- Table A3. Gray Fox records from Pelee Island, Ontario. Source: Judge and Haviernick (2002), NHIC data, and Bowman et al. (2013). Unconfirmed reports are not included.
- Table A4. Gray Fox records from the north shore of Lake Erie in Ontario. Source: Judge and Haviernick (2002) and updated using NHIC data. Unconfirmed reports are not included.
- Table A5. Gray Fox observations from the northeastern shore of Lake Ontario and St. Lawrence, Ontario. Judge and Haviernick (2002), updated using NHIC data.
- Table A6. Gray Fox records from Quebec. Source: MFFP, unpub. data.
- Table A7. Gray Fox records from New Brunswick.
- Appendix 2. Threats Calculator for Gray Fox.
COSEWIC
Committee on the Status
of Endangered Wildlife
in Canada

COSEPAC
Comité sur la situation
des espèces en péril
au Canada
COSEWIC status reports are working documents used in assigning the status of wildlife species suspected of being at risk. This report may be cited as follows:
COSEWIC. 2015. COSEWIC assessment and status report on the Gray Fox Urocyon cinereoargenteus in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. xii + 54 pp. (Species at Risk Public Registry website).
Previous report(s):
COSEWIC. 2002. COSEWIC assessment and update status report on the grey fox Urocyon cinereoargenteus interior in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. vi + 32 pp.
Judge, K.A., Haviernick, M. 2002. Update COSEWIC status report on the grey fox Urocyon cinereoargenteus in Canada, in COSEWIC assessment and update status report on the grey fox Urocyon cinereoargenteus interior in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 1-32 pp.
Steers, S. 1979. COSEWIC status report on the grey fox Urocyon cinereoargenteus in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 13 pp.
COSEWIC would like to acknowledge Jennie Pearce (Pearce & Associates Ecological Research) for writing the status report on the Gray Fox, Urocyon cinereoargenteus, in Canada, prepared under contract with Environment Canada. This report was overseen and edited by Graham Forbes, Co-chair of the COSEWIC Terrestrial Mammals Specialist Subcommittee.
For additional copies contact:
COSEWIC Secretariat
c/o Canadian Wildlife Service
Environment Canada
Ottawa, ON
K1A 0H3
Tel.: 819-938-4125
Fax: 819-938-3984
COSEWIC E-mail
COSEWIC Website
Également disponible en français sous le titre Évaluation et Rapport de situation du COSEPAC sur le Renard gris (Urocyon cinereoargenteus) au Canada.
Cover illustration/photo:
Gray Fox -- Photo: John Woodcock.
Gray Fox
Urocyon cinereoargenteus
The Gray Fox (Urocyon cinereoargenteus) has a typical fox-like appearance but the pelage is a grizzled grey colour, with reddish regions on the neck, sides, and legs. There is a prominent black stripe running the length of the back down to the tip of the tail. The Gray Fox is significant in the phylogeny of canids because it is considered to be the basal member of the Canid family. It is also the most arboreal of canids, and can partially rotate its ankle bones to facilitate descending trees.
The Gray Fox is generally found from south-central Canada to northern parts of South America and is expanding its range in the United States northward and eastward. They were present historically in southern Ontario but currently are likely breeding in two regions in Canada: Rainy River-Thunder Bay (hereafter; 'Northwestern Ontario'), and Pelee Island, Ontario. In this report, the Gray Fox in southeastern Manitoba, Quebec, and New Brunswick are considered to be extralimital or dispersers/visitors or, more appropriately, 'occasional' animals that have dispersed from adjacent populations in the United States and have not likely established breeding populations in these parts of Canada.
The Gray Fox's distribution is closely associated with deciduous forest. Gray Foxes den in many different kinds of substrates, usually located in dense brush close to a water source. Gray Foxes are considered habitat generalists and are partially tolerant of human disturbances, although they are generally more secretive than Red Fox (Vulpes vulpes), and so are seen less often.
Gray Fox are the most omnivorous of the North American canids and will consume vegetable matter, such as fruit, year-round. Gray Foxes in Canada are thought to breed from mid-February to mid-March, as they do in the northeastern United States. Most female Gray Foxes breed in their first year and have one litter of 3 - 4 kits per year. Gray Foxes are typically nocturnal or crepuscular. The basic social unit consists of an adult male and female and their offspring, and this group maintains a home range. The adult sex ratio is usually 1:1 and Gray Fox are assumed to be monogamous.
The population size is unknown. No quantitative population-level study of Gray Fox in Canada has been published but, based on the few records, it seems likely that the number of mature individuals present in Canada is fewer than 110. Populations of Gray Fox have been increasing in the northern US, concurrent with a northward range expansion that resulted in additional records in Canada.
Mortality from trapping likely is preventing the establishment of breeding sub-populations in much of the Canadian range outside Pelee Island. The most important factor affecting Gray Fox populations in the United States is trapping. The effect of harvest in Canada is unknown but most records in most sub-populations derive from incidental trapping. Because of the small population size, any significant mortality factor, such as high Coyote predation and diseases (including canine distemper and rabies), could become significant limiting factors. In the two sub-populations with evidence of breeding, but mainly in the Northwestern Ontario sub-population, the likely threats are mortality from trapping, and roadkill. The overall threat score was high.
NatureServe ranks the Gray Fox globally as very common (G5); however, in Canada it is ranked as Critically Imperilled (N1). The Gray Fox is listed as Threatened on Schedule 1 of the federal Species at Risk Act (SARA). In Ontario, the Gray Fox is listed as Threatened under the Ontario Endangered Species Act, 2007. In Quebec, the Gray Fox is considered an occasional resident and is not ranked or listed. In Manitoba, Quebec, and New Brunswick, Gray Foxes are considered to be occasional dispersers from the United States. The Gray Fox is thought to be present, although rare, in St. Lawrence Islands National Park, it has been found in Fish Point Provincial Nature Reserve on Pelee Island, and it is occasionally reported from the Whiteshell Provincial Park region in southeastern Manitoba.
| Summary Items | Information |
|---|---|
| Generation time (usually average age of parents in the population; indicate if another method of estimating generation time indicated in the IUCN guidelines (2008) is being used) Using IUCN Method 1 (average age of parents), with juvenile and adult survival rates to be 0.31 and 0.43 respectively, fecundity to be 3.7, and age at first breeding as year 0. Estimated survival rates are based on US populations subject to trapping. |
Probably 2 yrs |
| Is there an [inferred] continuing decline in number of mature individuals? Data on mature animals do not exist. However, in Northwestern Ontario, sightings may be increasing and the presence of mature females, a lactating female, and pairs indicates breeding likely is occurring; this population may be increasing. The population on Pelee Island may be stable. Records in southeastern Quebec are increasing but most of these likely are not mature animals. |
Unlikely |
| Estimated percent of continuing decline in total number of mature individuals within [5 years or 2 generations] (See previous) |
Unknown |
| [Suspected] percent [increase] in total number of mature individuals over the last [10 years, or 3 generations]. (See previous) |
Unknown |
| [Suspected] percent [increase] in total number of mature individuals over the next [10 years, or 3 generations]. Projection difficult because threats occur with unknown severity and limited data on mature animals exist. The Gray Fox sub-population in Northwestern Ontario is projected to increase while the sub-population on Pelee Island likely is stable. The number of dispersers into Manitoba, mainland Ontario, Quebec, and New Brunswick is likely to increase, and breeding may occur in the future. |
Unknown |
| [Suspected] percent [increase] in total number of mature individuals over any [10 years, or 3 generations] period, over a time period including both the past and the future. (See previous) |
Unknown |
| Are the causes of the decline clearly reversible and understood and ceased? Not applicable; sub-populations increasing and no evidence of decline in breeding (except for eastern Ontario where there were breeding records in the 1950s). |
N/A |
| Are there extreme fluctuations in number of mature individuals? Limited data on mature animals exists. |
Unknown |
| Summary Items | Information |
|---|---|
| Estimated extent of occurrence Based on minimum convex polygon method that includes only known or suspected areas with breeding in Pelee Island and Northwestern Ontario. The higher value includes areas with recent records (i.e., in last 20 years) of animals that have dispersed into Canada, but likely are not breeding. |
EOO: 35 173 km2 (625 517 km2) |
| Index of area of occupancy (IAO) (Always report 2x2 grid value). Based on possible breeding sites in Pelee Island and Northwestern Ontario. The higher value includes areas with recent records (i.e., in last 20 years) of animals that have dispersed into Canada, but likely are not breeding. |
IAO: 96 km2 (200 km2) |
| Is the population severely fragmented? The two breeding subpopulations are isolated from each other but the Northwestern Ontario sub-population is connected to the nearest US populations by dispersal. The extent of isolation of the Pelee Island population from other sub-populations is unknown. |
No |
| Number of locations (Note: See Definitions and Abbreviations on COSEWIC website and IUCN (Feb 2014) for more information on this term.) Pelee Island likely represents one location because animals exist within a relatively small area where threat of accidental trapping likely is similar, but the sub-population in Northwestern Ontario experiences trapping pressure that likely varies throughout the region. |
Unknown; likely > 10 |
| Is there an [projected] continuing decline in extent of occurrence? The EOO has increased with addition of records from Northwestern Ontario. |
No |
| Is there an [inferred] continuing decline in index of area of occupancy? IAO increased since 2002 with establishment of a breeding sub-population in Northwestern Ontario. |
No |
| Is there an [inferred] continuing decline in number of populations? The number of sub-populations has increased since 2002 with the establishment of a breeding sub-population in Northwestern Ontario. |
No |
| Is there an [inferred] continuing decline in number of locations*? See previous |
No |
| Is there an [inferred] continuing decline in [area, extent and/or quality] of habitat? No decline in habitat is evident. |
No |
| Are there extreme fluctuations in number of populations? The number of sub-populations with breeding has changed from 1 to 2 since 2002, but this is not considered extreme. |
No |
Are there extreme fluctuations in number of locations? (Note: See Definitions and Abbreviations on COSEWIC website and IUCN (Feb 2014) for more information on this term.) The number of locations with evidence of breeding has changed from 1 to 2 since 2002, but this is not considered extreme. |
No |
| Are there extreme fluctuations in extent of occurrence? The extent of occurrence has increased since 2002 through the inclusion of records from Northwestern Ontario; however, this is considered to be a range expansion and not a fluctuation related to risk of extirpation. |
No |
| Are there extreme fluctuations in index of area of occupancy? See above. | No |
| Summary Items | Information |
|---|---|
| Population | Unknown |
| Northwestern Ontario sub-population Limited data on mature individuals available. Probably fewer than 50 mature individuals (males and females); breeding is likely. | Unknown |
| Pelee Island, Ontario sub-population Limited data on mature individuals available. Probably fewer than 60 mature individuals; breeding confirmed. | Unknown |
| Manitoba/ parts of Ontario/Quebec / New Brunswick Animals are not included because most are likely sub-adult individuals dispersing from adjacent US populations and evidence of breeding is not strong enough, at present. | N/A |
| Total Estimated < 110 mature individuals | Unknown |
| Summary Items | Information |
|---|---|
| Probability of extinction in the wild is at least [20% within 20 years or 5 generations, or 10% within 100 years]. PVA not conducted |
N/A |
Mortality rates in Canada of Gray Fox dispersing from the United States likely are preventing the establishment of breeding populations and expansion in most sub-populations. Limiting factors include Coyote predation and disease.
| Summary Items | Information |
|---|---|
| Status of outside population(s)? Minnesota population expanding northward; Michigan, New York, Vermont, New Hampshire populations stable; Maine population expanding northwestward. The population in Ohio near Pelee Island is declining. |
US populations increasing in most adjacent states |
| Is immigration known or possible? Animals in northwestern Ontario and Quebec sub-populations are assumed to be supported by immigration from US. Immigration to the Pelee Island sub-population from US is unknown, but possible from mainland in winter. Individuals observed in Manitoba, Quebec, and New Brunswick likely emigrated from the US. |
Yes |
| Would immigrants be adapted to survive in Canada? Environmental conditions assumed to be similar |
Yes |
| Is there sufficient habitat for immigrants in Canada? | Yes |
| Is rescue from outside populations likely? Within the EOO, rescue in the Northwestern Ontario sub-population is likely; rescue to Pelee Island sub-population is unknown. |
Partially |
| Summary Items | Information |
|---|---|
| Is this a data sensitive species? | No |
Designated Special Concern in April 1979. Status re-examined and designated Threatened in May 2002 and November 2015.
Since the publication of the 2002 COSEWIC status report, a second sub-population containing breeding animals has established, as well as increased presence of Gray Fox in Quebec and New Brunswick. Gray Fox in Manitoba continue to be present though breeding has not been established. Individual animals recorded in New Brunswick, Quebec, and Manitoba are considered to be individuals that have recently dispersed from adjacent populations in the United States. Because of the absence of breeding evidence, animals in New Brunswick, Quebec, and Manitoba are not considered relevant to status.
As part of COSEWIC status assessments, Aboriginal Traditional Knowledge (ATK) reports are prepared by the Aboriginal Traditional Knowledge subcommittee (ATK SC). These initial reports compile and summarize ATK relevant to status assessment when ATK information is available and readily accessible. A Gathering Report may be undertaken if there are significant knowledge gaps, or if major contradictions exist between ATK and other forms of knowledge. The ATK SC has not identified ATK specific to the Gray Fox in Canada.
COSEWIC would like to acknowledge Jennie Pearce for writing the status report on the Gray Fox, prepared under contract with Environment Canada. Modifications to the status report after acceptance of the provisional report were overseen by Graham Forbes, Co-chair of the COSEWIC Terrestrial Mammals Specialist Subcommittee (TM SSC), based on comments from jurisdictions, the TM SSC, COSEWIC members, and external experts.
The Committee on the Status of Endangered Wildlife in Canada (COSEWIC) was created in 1977 as a result of a recommendation at the Federal-Provincial Wildlife Conference held in 1976. It arose from the need for a single, official, scientifically sound, national listing of wildlife species at risk. In 1978, COSEWIC designated its first species and produced its first list of Canadian species at risk. Species designated at meetings of the full committee are added to the list. On June 5, 2003, the Species at Risk Act (SARA) was proclaimed. SARA establishes COSEWIC as an advisory body ensuring that species will continue to be assessed under a rigorous and independent scientific process.
The Committee on the Status of Endangered Wildlife in Canada (COSEWIC) assesses the national status of wild species, subspecies, varieties, or other designatable units that are considered to be at risk in Canada. Designations are made on native species for the following taxonomic groups: mammals, birds, reptiles, amphibians, fishes, arthropods, molluscs, vascular plants, mosses, and lichens.
COSEWIC comprises members from each provincial and territorial government wildlife agency, four federal entities (Canadian Wildlife Service, Parks Canada Agency, Department of Fisheries and Oceans, and the Federal Biodiversity Information Partnership, chaired by the Canadian Museum of Nature), three non-government science members and the co-chairs of the species specialist subcommittees and the Aboriginal Traditional Knowledge subcommittee. The Committee meets to consider status reports on candidate species.
The Canadian Wildlife Service, Environment and Climate Change Canada, provides full administrative and financial support to the COSEWIC Secretariat.
Common Name: Gray Fox (Wilson and Reeder 2005), Grey Fox (Canadian English), Renard gris (French)
Aboriginal Names:no name used by First Nations within Canadian range identified
Class: Mammalia
Order: Carnivora
Family: Canidae
Genus: Urocyon
Species: cinereoargenteus Schreber (1775)
Eight subspecies are recognized in North America (Fritzell and Haroldson 1982; Wilson and Reeder 2005), with three subspecies believed to occur in Canada: U. c. borealis Merriam, U. c. cinereoargenteus (Schreber) and U. c. ocythous Bangs (Banfield 1974) (Figure 2). Seven additional subspecies are recognized in Central and South America (Fritzell 1987). The validity of subspecific designations is discussed in the Population Spatial Structure and Variability, and Designatable Units sections.
The Gray Fox (Figure 1) is a member of the family Canidae, a group that includes dogs, wolves, foxes, and jackals. Gray Fox are similar in size and appearance to the Red Fox (Vulpes vulpes). The Gray Fox has a dark longitudinal stripe, made up of coarse black-tipped hairs, running the length of its back and tail (Fritzell and Haroldson 1982). The sides and back of its pelage are grizzled grey, and the underfur is buffy. There is white fur on the ears, face, throat, chest, belly and hind legs but the tail is black-tipped, in contrast to the white tip of Red Fox. There is a distinctive black muzzle patch in front of each eye and on the lower jaw. Compared to the Red Fox, Gray Fox have shorter legs, a shorter muzzle, rounder footprints, larger toe pads, and more curved claws (Banfield 1974). Body length with tail is 136 cm (range: 107.5 - 156.8) and average weight for males is 4.1 kg, and 3.9 kg for females (Banfield 1974).

Long description for Figure 1
Photo of a Gray Fox, lateral view with head facing camera. A dark longitudinal stripe, made up of coarse black-tipped hairs, runs the length of its back and tail. The sides and back of its pelage are grizzled grey, and the underfur is buffy. There is white fur on the ears, face, throat, chest, belly, and hind legs, but the tail is black-tipped. There is a distinctive black muzzle patch in front of each eye and on the lower jaw.
Little information exists on the spatial structure for Gray Fox in the US, and none exists for the Canadian population. The subspecies designations for all Gray Fox were established in the early 1900s by comparing the morphometrics (i.e., skull, teeth, and body dimensions) and pelage of new specimens against the known type specimen. The first type specimen/subspecies was described from the Carolinas in 1775 and eventually named Urocyon cinereoargenteus cinereoargenteus (Schreber 1775; Hall 1981). The ocythous subspecies was based on a specimen from Wisconsin that had a larger skull and body, and more yellow pelage (Bangs 1899), and the borealis subspecies was based on a specimen from New Hampshire that was also larger, particularly in the dimensions of various teeth (Merriam 1903). The range of each subspecies was finalized by Hall (1981) who compared museum specimens and delineated boundaries by connecting locations of specimens along the range margins.
According to Hall (1981), Gray Fox in Manitoba and northwestern Ontario are designated as U. c. ocythous, the subspecies present in Minnesota. Gray Fox found in the Lake Erie region are believed to be U. c. cinereoargenteus, and Gray Fox in eastern Ontario, southern Quebec, and presumably the records for New Brunswick, are U. c. borealis (Figure 2). Gray Fox on the north shore of Lake Erie in Ontario (and on Pelee Island) are designated as U. c. cinereoargenteus,based on a marginal specimen record from Oxford County (Hall 1981), and that the population may receive cinereoargenteus subspecies originating from adjacent Michigan, Ohio and eastern New York.
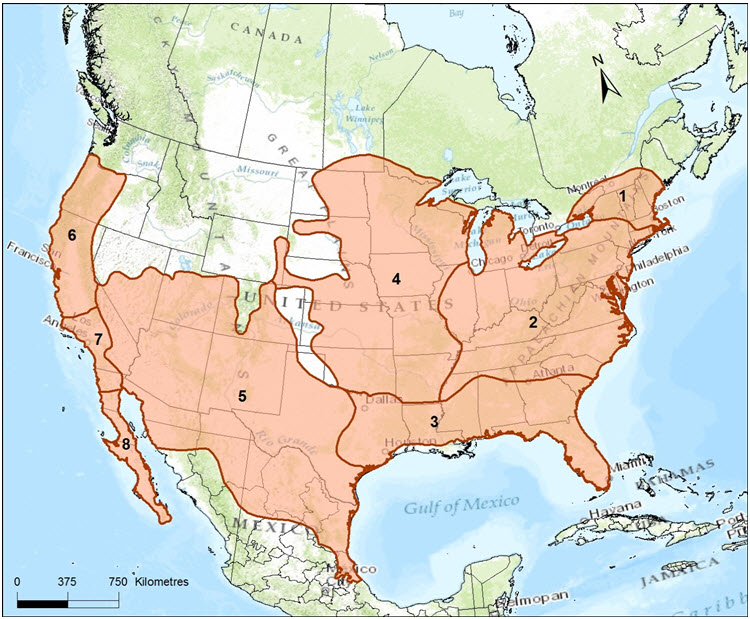
Long description for Figure 2
Map of the distribution of the Gray Fox in mainland North America, indicating locations of 8 of 15 subspecies. The Gray Fox has a wide distribution, from the Canadian border at Manitoba to Quebec, and southward through the eastern and southern United States, and to northern Colombia and Venezuela. The Gray Fox is absent from the northwestern United States and the Great Plains in the United States
A single study (Bozarth et al. 2011) addressed the genetic structure of three subspecies in eastern North America (borealis, cinereoargenteus, and floridus), two of which (borealis, cinereoargenteus) extend into Canada. No samples from Canada were included. Based on a sample of 303 Gray Fox, Bozarth et al. (2011) found no clear pattern of genetic structuring of Gray Fox haplotypes across the eastern United States. A lack of structuring suggests that the subspecies designations may not be valid (Bozarth et al. 2011). Similar results have been recorded in other mammals that survived in refugia during glaciation, then expanded northward (e.g., Wooding and Ward 1997; Aubry et al. 2009).
A rigorous statistical-based review of morphometric differences, or genetic differences, has not been conducted on Gray Fox. Such reviews, conducted on other carnivore species with strong dispersal abilities, have resulted in a much reduced number of North American subspecies; for example, Red Fox (10 subspecies, reduced to 2 genetic clades, Hall 1981; Kutschera et al. 2013), Gray Wolf (Canis lupus) (24 to 5 subspecies, Hall 1981; Nowak 1995), and Brown Bear (Ursus arctos) (86 to 7, or 2, subspecies, Merriam 1918; Rausch 1963; Hall 1984). It is likely that the number of subspecific designations of Gray Fox in eastern North America would be similarly reduced. Since the glacial period, there has been no apparent barrier in eastern-central North America to Gray Fox movement that would isolate populations and facilitate genetic uniqueness. The differences in body size have been the main criteria for subspecies but the differences are clinal, with dimensions increasing from south to north (Hall 1981), which is a common trait among mammals in North America (e.g. Ashton et al. 2000) and not a strong basis for designating subspecies (James 2010; Ennen et al. 2014).
The only genetic evidence for differences within Gray Fox is that samples from the northeast US have 4 haplotypes, compared to 36 in southward populations (Bozarth et al. 2011). However, it is believed that the haplotype difference likely resulted from a small founder population expanding northward from the refugium, rather than being the result of deep genetic divergence created by adapting in glacial refugia (Bozarth et al. 2011). Also, the genetic difference did not correspond to the subspecific boundaries; haplotype frequencies of borealis samples from Vermont, New Hampshire, and Maine were similar to cinereoargenteus samples from New York. Only a single sample (northern Ohio) was examined from the remaining Great Lakes states and consequently no conclusions can be drawn regarding the genetic distinctiveness of cinereoargenteus entering southwestern Ontario, or of ocythous entering Rainy River - Thunder Bay (hereafter, 'Northwestern Ontario) and Manitoba. However, given the species' dispersal abilities and lack of barriers in eastern North America, it is unlikely that Gray Fox in the Great Lakes region originated from a glacial refugium separate from the one used by southeastern Gray Fox, and which has not produced genetic structuring (Bozarth et al. 2011). Deep genetic divergence may have occurred for Gray Fox in western mountainous regions of the US but there is no evidence these animals have entered Canada.
A single designatable unit is recognized, based on the lack of genetic structuring, the mismatch between genetic and morphometric information used to define subspecies, and a general lack of rigour in historical subspecific designations in Gray Fox (see Population Spatial Structure and Variability). The areas containing breeding evidence are considered to be sub-populations because they are presently isolated and widely spaced within Canada.
The Gray Fox is considered the most basal member of the Canidae family, suggesting a North American origin of Canids 10 million years ago (Lindblad-Toh et al. 2005). Although abundance in Canada likely is too low to permit Gray Fox to affect ecosystems, where abundant in the United States, Gray Fox contribute to regulating small mammal abundance. The high level of frugivory in Gray Foxes suggests they may also play a role in seed dispersal.
In the United States, the Gray Fox has some economic importance as a furbearer but harvest is not permitted in Canada.
The Gray Fox has a wide distribution, from the Canadian border at Manitoba to Quebec, and southward through the eastern and southern United States, and to northern Colombia and Venezuela. The Gray Fox is absent from the northwestern United States and the Great Plains in the United States (Fritzell and Haroldson 1982; Figure 2).
In Canada, the Gray Fox has been recorded in Alberta, Manitoba, Ontario, Quebec, and New Brunswick (Figure 3; Table A1 - A7). Breeding populations appear to be limited to two parts of Ontario.

Long description for Figure 3
Map indicating the extent of occurrence (EOO) of the Gray Fox in Manitoba, Ontario, Quebec, and New Brunswick, based on recent records (within the last 20 years). Symbols indicate the locations of recent and historical records. Gray Fox have a total EOO of 625,517 square km within Canada's extent of jurisdiction.
A single Gray Fox was trapped near Lake Athabasca in Alberta in January 1950, over 2000 km from the nearest Gray Fox population (Moore 1952). Alberta is not included as part of the Canadian range of Gray Fox because this specimen is considered to be an extra-limital record, or vagrant (Moore 1952; Fritzell and Haroldson 1982; Smith 1993) and even perhaps an escaped captive (Fritzell 1987).
The first confirmed record of Gray Fox in Manitoba was an adult male trapped in 1957 near St. Adolphe, although an unconfirmed report also exists for Sprague from 1946 - 1947 (Sutton 1958). There have been < 10 confirmed, and numerous unconfirmed, records, all in southeastern Manitoba, primarily near Sprague, Whiteshell Provincial Park, St. Adolphe and near Hadashville (Table A1). The 2002 COSEWIC report cites a personal communication of I. McKay, a provincial furbearer biologist, that six to seven animals were harvested annually (dates not provided) from the Whiteshell Park area (Judge and Haviernick 2002). No records of these animals are available, and there is only one confirmed record from Whiteshell Park, in 2002. Fur harvest records from 1978 to 2014 do not contain Gray Fox (Manitoba Wildlife Branch 2015). Such a high number of past captures is of interest but further information on confirmed harvest, and breeding condition is lacking; there is no evidence of regularly occurring Gray Fox in Whiteshell Park presently (Berezanski pers. comm. 2014). Gray Fox continue to be intermittently reported in southeastern Manitoba, primarily as roadkill, during winter (Berezanski pers. comm. 2014). The records are all < 100 km, and mostly < 60 km, from known range in Minnesota. Breeding has not been confirmed in Manitoba, and the few individuals recorded since 2000 likely represent dispersing individuals from Minnesota (Berezanski pers. comm. 2014). The Manitoba government does not consider Gray Fox to be a breeding resident of the province.
Although Gray Fox were recorded before European colonization (see Fluctuations and Trends), in the last 50 years they have been recorded from only four areas: (1) the Rainy River - Thunder Bay area ('Northwestern Ontario' = northwest of Lake Superior bordering the US); (2) Pelee Island; (3) the north shore of Lake Erie from Windsor to Niagara Falls; and (4) the northeastern shore of Lake Ontario and St Lawrence. Despite Gray Fox being found around Sault Sainte Marie, Michigan (Rollings pers. comm. 2014), Gray Fox have not been recorded from adjacent Sault Ste. Marie, Ontario (Siccoly pers. comm. 2014).
(1) Northwestern Ontario (Table A2):
Gray Fox were first recorded in 1944 in Rainy River district (Banfield 1974) and have since been observed from Rainy River to Dorion, Ontario, with 25 to 28 confirmed Gray Fox individuals since 1944 (Judge and Haviernick 2002; Van den Broeck 2014a). Sightings appear to have increased since 2006 (Van den Broeck 2014a). In early March 2014, a mature male (4.0 kg) and mature female (4.2 kg) Gray Fox travelling together were killed by a vehicle near Rainy River (Van den Broeck 2014b). Subsequent necropsy revealed no evidence of breeding (Sadowski, pers. comm. 2015). Assessment of stomach contents for this pair revealed that the male stomach had a small quantity (25 ml) of crab apple fruit (Malus sp.) and cracked corn, while the female had an empty stomach with a few sunflower seed hulls in the lower intestine. Both foxes were thin with little body fat (Sadowski pers. comm. 2015). Also, a single Gray Fox was observed in this same general area in November 2013 (Van den Broeck 2014b). In 2010, two individuals were observed together near Thunder Bay, and in 2013, four individuals were observed together in the same general area (Van den Broeck 2014a). In summer 2015, a lactating female was repeatedly observed (and photographed July 1) in the Mountain Road area south of Thunder Bay, which is inferred to be the strongest evidence that a breeding population exists in this region (Deschamps pers. comm. 2015).
(2) Pelee Island (table A3):
The first confirmed record of Gray Fox on Pelee Island is in 1983 but it is not clear when they first arrived on the island. Breeding on Pelee Island was confirmed in 1 June 1998 when a 6-week old kit was observed (London Free Press 1998); additional unconfirmed observations of Gray Foxes denning under a house were reported in late May 2011 (M. McFarlane, NCC unpubl. data). A mature female (5.7 kg) road-killed fox collected in December 2012 on Pelee Island was found to have placental scars, indicating previous breeding. This fox was in good condition, with extensive fat deposits and 178 g of stomach contents, including mice, a small songbird, and other unidentified items (Sadowski pers. comm. 2015). In 2013, sightings of Gray Fox were primarily obtained from the northern section of Pelee Island (Bowman et al. 2013; McFarlane 2013).
(3) North Shore of Lake Erie (southwestern Ontario) (table A4):
Based on the cluster of records in the Niagara Falls area, it is believed that Gray Fox entered from western New York in 1948 (Peterson et al. 1953; Banfield 1974). Between 1949 and 2002, Gray Fox have been reported from the north shore of Lake Erie north to Bruce Peninsula, and as far east as Toronto. A total of 42 - 45 confirmed sightings have been recorded in this region, but none since 2002
(4) Northeastern Shore of Lake Ontario and St. Lawrence (table A5):
The first record was of an individual crossing the St. Lawrence River in 1942 (Banfield 1974). Peterson et al. (1953) cited a news release that Gray Fox were breeding in the general area of Leeds, Grenville, Stormont, Dundas, and Glengarry counties in 1952, with one or two Gray Fox shot annually. A total of 19 confirmed sightings have been reported between 1942 and 2014 (Judge and Haviernick 2002), but there are no breeding records since the 1950s. An unconfirmed report of a grey-colored fox high up in a cedar (Thuja sp.) tree near Minto was reported in June 2009 (Linnitt pers. comm. 2015). A road-killed fox was collected on the St. Lawrence Islands Parkway in October, 2008. The sex of this fox could not be determined due to carcass condition, but the fox appeared to be a juvenile, based on tooth wear (Bruce pers. comm. 2015). An unconfirmed report of a grey-colored fox with a black tip on its tail frequenting a heavily wooded area near Cavendish Lake was reported in September 2009 (Arscott pers. comm. 2015). The fox was observed on several evenings crossing a small gravel road between a clearing and the woods. Two foxes were observed in 2014, one 28km northwest of Smith Falls, and the other, 23 km east of Smith Falls.
Thirty-six confirmed Gray Fox specimens have been recorded since 1893/1894, from four areas in Quebec (MFFP unpub. data; Figure 3): (1) southwestern Montérégie region, (2) southwestern Estrie region, (3) southeastern Estrie region, and (4) southeastern Chaudière-Appalaches region. There have been 30 records made in the last 20 years (1996-2015).
(1) Southwestern Montérégie Region (table A6):
Twenty confirmed sightings have been recorded from the southwestern Montérégie region between 1975 and 2014, all within 42 km of the New York border. In the 1990s, one male and one female were trapped near St-Anicet within a three-week interval (Jolicoeur et al. 2011). Ten of the 20 sightings have been reported within the last 3 years, predominantly from the Hemmingford and Saint-Bernard-de-Lacolle area, which is within 6 km of the New York/Vermont border.
(2) Southwestern Estrie Region (table A6):
The first record of a Gray Fox in Canada was of an individual from Compton County near Johnville in the winter of 1893/1894 (Anderson (1939, cited in Judge and Haviernick 2002)). Four additional sightings were reported in winter between 2011 and 2013, including a juvenile female in 2011, within 45 km of the Vermont/New Hampshire borders.
(3) Southeastern Estrie Region (table A6):
Four confirmed records have been reported from this region in winter. The first individual was trapped in the 1940s, and the remaining three individuals were killed or trapped between 2011 and 2013, within 15 km of the Maine border.
(4) Southeastern Chaudière-appalaches Region (table A6):
Five Gray Fox have been trapped in this region between 1986 and 2007, within 45 km of the Maine border.
The number of recent records in Quebec, particularly within southwestern Montérégie and western Estrie, suggests Gray Fox may be resident in Quebec but evidence of breeding is weak. Of the 30 records from the last 20 years, 12 animals were identified to age or gender, with 9 males and 3 females, of which 4 were considered adult males, and 1 was a juvenile female (Table A6). Lactating females or kits have not been recorded. All records are within 50 km of known, or suspected, breeding populations in the United States. One scenario is that mortality rates from incidental trapping is preventing a breeding population from establishing. The strongest evidence of breeding was of a juvenile female killed near Sherbrooke on December 12, 2011 (Table A6). Dispersal likely occurs in October (see Dispersal), and it is possible this animal was born in Vermont and dispersed to Quebec over the next few months. Alternatively, the animal may have been born in Quebec because females generally disperse less frequently than males (see Dispersal), and the 50 km is a considerable distance. At present, the weight of evidence is that Gray Fox are most likely dispersing into Quebec from New York, Vermont, and Maine, and therefore Gray Fox in Quebec are considered to be part of the Canadian range, but are not relevant to status. It seems probable that breeding will be recorded in Quebec in the near future.
Two Gray Fox have been recorded in New Brunswick (Table A7). In October 2007, a single, sub-adult male Gray Fox was trapped near Rollingdam, Charlotte County, New Brunswick (McAlpine et al. 2008). In September 2014, a Gray Fox was observed using a motion-activated infra-red camera at the north end of Oromocto Lake, York County, NB (McAlpine et al. submitted). Both individuals are believed to have dispersed from Maine because no other records exist in the province, and the nearest records (Bangor, Maine) are over 130 km distant. The records are included in the extent of occurrence calculation but it is likely premature to conclude that NB is part of Gray Fox range. Unlike in Quebec or Manitoba, where a resident population exists near the Canadian border, there is a 100 km gap between the NB border and the range in Maine. There is no evidence of a breeding population in New Brunswick.
Under COSEWIC guidelines, a vagrant is a wildlife species whose geographical range is outside Canadian jurisdiction and that has never established a viable population in Canada, but may occasionally be recorded in Canada (Appendix C, Operations and Procedures Manual). Species that lack a regularity of occurrence, such as vagrants, are not considered valid for assessment (Appendices E3, F9, Operations and Procedures Manual). Under IUCN guidelines, the term 'visitor' is used for a taxon that does not reproduce within a region but regularly occurs within its boundaries either now, or during some period of the last century (IUCN 2010). However, the vagrant or visitor concept does not apply well to the Gray Fox situation in Canada. Gray Fox are established as a breeding, resident species in two areas of Canada and the species is eligible for assessment, but most of the distribution is in areas composed of apparently non-breeding (not mature) animals that have dispersed into Canada from the United States. Evidence of breeding is confined to Pelee Island, and Northwestern Ontario. Gray Fox that were recorded in Alberta, Manitoba, southeastern Ontario, Quebec, and New Brunswick are considered to represent irregular or occasional dispersing animals that presently are not part of an established breeding population. As such, these records are considered to be part of the extent of occurrence in Canada but are not considered significant to the viability of Gray Fox in Canada and therefore are not included in the population estimate.
The extent of occurrence (EOO) and index of occupancy (IAO) is calculated for both the areas where breeding is likely, and the total area of records in Canada (see Canadian Range). Based on sightings within the last 20 years (i.e., 1994 - 2015, an arbitrary value but one that likely captures recent climatic and habitat factors that influence Gray Fox distribution - see Habitat Trends, Fluctuations and Trends), Gray Fox have a total extent of occurrence of 625,517 km2, and an IAO (2km x 2km grid) of 200 km2. Based on all recent records (i.e., in the last 20 years) for the two sub-populations with evidence of breeding activity (Pelee Island and northwestern Ontario), the EOO is 35,173 km2, and the IAO is 96 km2 (Figure 3).
Data were obtained from provincial Conservation Data Centres, public reports of Gray Fox observations, trapping records, and U.S. state wildlife departments. A systematic camera-trapping survey was undertaken during 2012 - 2013 on Pelee Island (Bowman et al. 2013). Otherwise, most data are based on voluntary public reports of sightings, road-kills, and incidental trapping.
Data on population size and trends in the United States are primarily based on hunting and trapping data. In Quebec and Ontario, hunters and trappers that kill Gray Fox are required to report this information and submit the carcass if requested to do so; if the animal is incidentally captured and released there is no requirement to report the capture. Elsewhere in Canada, there is no specific requirement for hunters and trappers to report incidental capture of the Gray Fox.
Habitat use by Gray Fox in Canada is poorly known. Researchers with the Ontario Ministry of Natural Resources, Rabies Research Unit, radio-collared a single adult male Gray Fox and radio-tracked it from October 1980 to November 1981 in Lambton County, Ontario, an area of cultivated fields interspersed with deciduous woodlots dominated by Sugar Maple (Acer saccharum) within a network of roads (Bachmann and Lintack 1982). The Gray Fox male was located most often within woodlots and moved most often within 100 m of the woodlot edge. The male only occasionally moved into open, less wooded areas, or crossed roads (Bachmann and Lintack 1982). All observations of Gray Fox in the Pelee Island camera survey occurred in deciduous forests (Bowman et al. 2013; Bowman pers. comm. 2014). These are the only two studies of Gray Fox habitat use in Canada.
Habitat use determined from northeastern US range likely applies to Gray Fox in Canada. Gray Foxes use a greater proportion of wooded habitat than other North American fox species, and brush and tree cover are important habitat components (Cypher 2003). In eastern North America, Gray Foxes prefer a mix of fields and woods, rather than large tracts of homogenous forest (Wood et al. 1958). In Wisconsin, areas interspersed with forest and farmland are considered high-quality habitat (Richards and Hines 1953; Petersen et al. 1977). In Illinois, Gray Foxes use old fields more than expected based on availability, and agricultural and brushy habitat less than expected, and woodlands in proportion to their availability (Follmann 1973). Old fields appear to be used for foraging (Follmann 1973; Yearsley and Samuel 1980), whereas woodlands, particularly dense or brushy stands, commonly are used for daytime cover (Follmann 1973; Yearsley and Samuel 1980; Hallberg and Trapp 1984; Haroldson and Fritzell 1984).
In their northern US range, Gray Foxes use dens primarily during parturition and pup rearing, although they may occasionally use dens at other times to avoid predators (Cypher 2003). Dens may be dug or modified burrows of other species (Schmeltz and Whitaker 1977). They will also den in wood piles, brush piles, rock crevices, hollow logs and trees, hollows under shrubs, and under abandoned houses (Latham 1943; Sullivan 1956; Trapp and Hallberg 1975; Fritzell and Haroldson 1982). Gray Fox have been observed denning in hollow trees 7.6 m (Grinnell et al. 1937) to 9.1 m (Davis 1960) above the ground. In eastern deciduous forests, dens typically face east, southeast or south (Richards and Hines 1953) and are located in brushy or wooded areas (Layne and McKeon 1956a; Nicholson et al. 1985).
Daytime resting sites are typically located in dense vegetation (Hallberg and Trapp 1984; Nicholson et al. 1985), but they have also been observed resting in the shade of boulders (Trapp 1978). In Canada, Bachmann and Lintack (1982) found a single male used at least three rest sites, all of which were within brush piles. With the exception of one site in a fallow field, all resting sites observed were in woodlots within 50 m of the field edge.
The unique climbing behaviour of Gray Fox is summarized in Fritzell and Haroldson (1982). Gray Fox exhibit adaptations that facilitate tree climbing, such as a stocky build, relatively short legs with long sharp re-curved claws, and an ability to rotate their foreleg more than other canids, which may help grip better when climbing trees. Gray Foxes climb trees and shrubs to forage, rest, and escape predators. They can climb vertical, branchless tree trunks to a height of 18 m by clasping the trunk with their forelegs and pushing upwards with their hind legs. Most waterways are unlikely to be barriers to dispersal because Gray Fox are capable swimmers (Cypher 2003).
Coarse-scale vegetation in northwestern Ontario is primarily deciduous, mixed or coniferous forest used for timber production, except in the Agassiz Plains area in the northwestern corner that contains approximately 31% agricultural lands mixed with forest (Ontario Partners in Flight 2008a). Animals in adjacent Manitoba exist in forest cover similar to the Agassiz Plains, but with a high proportion of grassland and agricultural land, mixed with forest. The availability of Gray Fox habitat within northwestern Ontario and southeastern Manitoba is likely stable.
Pelee Island has approximately 20% forest cover (Essex Region Conservation Authority 2011). Most of the larger forest patches on Pelee Island are contained within protected areas, and as a result, the availability of Gray Fox habitat within Pelee Island is likely stable.
In southern Ontario, loss of forested habitat was substantial from the early part of the 19th century, as forest was converted to agricultural production. Today, land cover in southwestern Ontario and the northeastern shore of Lake Ontario comprises approximately 75% crops, 4% fields and 14% forest (Ontario Partners in Flight 2008b). Forest cover is predominantly restricted to woodlots within an agricultural matrix; total forest cover is stable, or may be slightly increasing with reforestation efforts (Ontario Partners in Flight 2008b). Forest cover is lowest in Essex Region in extreme southwestern Ontario and highest in the northern parts of the Gray Fox range along the north shore of Lake Erie and Lake Ontario. In Essex Region, approximately 77% of the land base is used for agriculture, with cash-crop farms, specialty crops, orchards and greenhouse farming being the most prevalent agricultural uses. The remainder of the area is 19% urban land use and approximately 6% forest. Similar patterns of shrub and tree cover are found in Niagara Region (Niagara Region Conservation Authority 2013), where the greatest abundance of forest cover is along the Niagara Escarpment and in the southeast of the region (Niagara Region Conservation Authority 2013). While the availability of Gray Fox habitat (brushy/forested areas interspersed with fields) is anticipated to be stable over time, high intensity agricultural landscapes do not provide optimal habitat for Gray Foxes, possibly because there is insufficient brush and tree cover for shelter. The Essex region is the closest Canadian mainland to Pelee Island, so the rarity of forest cover in Essex region may limit opportunities for emigration from Pelee Island to the mainland.
Habitat in southern townships of Quebec is primarily a mixture of agricultural areas, fields, and forest. Forests of young to mature age exist over much of southern New Brunswick where the two Gray Fox were recorded. Because Gray Foxes prefer a mix of fields and woods, and they will also use sub-urban habitats, it is likely that vegetative habitat suitable to support Gray Foxes is available in southern Quebec and southwestern New Brunswick.
Very little research has been conducted on Gray Foxes range-wide, and consequently basic ecological and demographic information is still needed. Information summarized here is based mainly on studies conducted in the United States.
All North American foxes are opportunistic foragers, with the Gray Fox perhaps the most omnivorous of all species (Cypher 2003). Gray Foxes eat medium to small-sized mammals, carrion, birds, insects such as Orthoptera, Coleoptera, and Lepidoptera, fruit, corn, nuts, and grains (Fritzell and Haroldson 1982; Cypher 2003). Recent necropsies of road-killed Gray Fox collected in Ontario reveal a range of food items including crab apples, sunflower seeds, mice, a songbird, and corn (Sadowski pers. comm. 2015).
Gray Foxes show seasonal changes in diet, with rabbits and small mammals forming the larger component of the diet in winter (Hatfield 1939; Fritzell 1987), insects (particularly Orthoptera) dominating stomach contents in summer, and fruit and nuts becoming an increasingly dominant part of the diet in the autumn (Pils and Klimstra 1975). Vegetable matter, such as fruit, is an important year-round diet component.
The sex ratio and age structure in Gray Fox populations is difficult to determine because most estimates are based on trapping data, which may be biased toward juvenile male capture (Fritzell 1987). Davis and Wood (1959) reported that 48 - 61% of Gray Fox harvest consists of individuals less than one year of age. In Wisconsin, 60 - 72% of females captured over a 3-year period were <1 year old (Richards and Hine 1953). In Alabama, 7% of foxes were juvenile (Sullivan and Haugen 1956). In Wisconsin, 66% of Gray Foxes harvested were juvenile (<1 year), 16% were yearling (1.5 years) and 15% were adults (Root and Payne 1985). In the single study of a non-harvested population, Weston and Brisbin (2003) found a gender bias towards females (59%), and 56% of animals were adults during a 50-year period in South Carolina.
Gray Foxes were assumed to be monogamous but more recent research identified multiple paternal sources in up to 57% of the litters (Weston Glenn et al. 2009). Unmated individuals locate mates in autumn, with courtship taking place in early winter (Fritzell 1987). Male behaviour during pup-rearing is not well understood, but their presence near the den during pup-rearing suggests that they contribute (Chamberlain and Leopold 2000). Nicholson et al. (1985) concluded that females primarily provision the pups.
The breeding season varies across the geographic range of the Gray Fox, with northern populations breeding later than southern populations (Trapp and Hallberg 1975; Fritzell and Haroldson 1982; Fritzell 1987). In New York and Wisconsin, peak breeding activity occurs from mid-February to mid-March (Sheldon 1949; Richards and Hine 1953; Layne and McKeon 1956b; Jackson 1961; Root 1981). The gestation period is 53 - 63 days (Fritzell 1987). The average litter size of 3.7 was observed in eight studies that counted in utero embryos in pregnant females or placental scars in lactating females. From 1 to 9 fetuses or scars were recorded, with 4 being the most common (Fritzell and Haroldson 1982; Root and Payne 1985). Fetal and neonatal sex ratios are not significantly different from 1:1 (Layne 1958). Juveniles reach adult size / weight at approximately 210 days (Wood 1958). Male and female Gray Fox become sexually mature at approximately 10 months of age (Follman 1978; Root 1981) and most breed in their first year (Root and Payne 1985).
No information is available on survival rates of Gray Foxes in Canada, and information from the United States is generally based on trapping mortality (Fritzell and Haroldson 1982; Cypher 2003). Within harvested populations, juvenile mortality may be high. In Florida, Lord (1961) estimated that 43 - 47% of Gray Foxes died during their first seven months and that mortality rates for adults were 61 - 64%. In southern Georgia, 50% of juveniles died by the end of their first summer and 90% did not survive their first winter; annual adult mortality rates were 50% (Wood 1958). In an area protected from trapping harvest, the annual adult mortality rate was 31% (Weston and Brisbin 2003). The high level of harvest pressure has been maintained in much of the US range, suggesting that Gray Fox are relatively productive (Fritzell 1987; Weston and Brisbin 2003).
Information on longevity in Gray Fox is obtained from populations subject to trapping mortality. These data suggest most Gray Foxes do not live past 4-5 years of age (Wood 1958; Lord 1961). The single Gray Fox radio-tracked in Ontario was estimated to be 4.5 years old (Backmann and Lintack 1982). Estimated generation time is 2 years, based on estimation Method 1 (average age of parents) of the IUCN guidelines (IUCN 2014). In this calculation, juvenile, and adult, survival rates were estimated as 0.31, and 0.43, respectively, fecundity as 3.7, and age at first reproduction as 0 years.
The Gray Fox is believed to be a southern, warm-adapted species (Waters 1964), and the northern edge of the species range may be defined by the species' ability to tolerate colder temperatures (see Fluctuations and Trends). Gray Foxes are generally tolerant of humans, being found on the outskirts of cities in the United States and they have been reported around bird feeders in northwestern Ontario. Human-altered landscapes that provide a mix of forest and fields create suitable habitat for the Gray Fox (see Habitat Use).
Gray Foxes are primarily nocturnal or crepuscular for foraging (Fritzell and Haroldson 1982), with resting during the day. During the day some movement between resting places is common (Trapp 1978). During its nightly activities, the adult male Gray Fox radio-tracked in Ontario was often found making circuits of the woodlots within its home range (Bachmann and Lintack 1982). On Pelee Island, nearly all Gray Fox detected on wildlife cameras in 2012/2013 were captured between 20:00 and 09:00, with peak activity between 22:00 and 01:00 (McFarlane, unpub. data 2013). One Gray Fox was captured on camera during the day (at 14:35, McFarlane unpubl. data).
In Mississippi, Gray Foxes moved at greater rates during pup-rearing (446 ± 85 m/hr) and winter (432 ± 83 m/hour, mean + SE) than during the breeding period (344 ± 46m/hour) (n=37, Chamberlain and Leopold 2000). They moved further during crepuscular (454 ± 69 m/hr) and nocturnal periods (443±64 m/hr) than during diurnal periods (265 ± 67 m/hr).
The basic social unit consists of an adult male and female, and their offspring, and this group maintains a home range that is largely exclusive of other such groups. It is unclear whether Gray Foxes are territorial. Gray Fox routinely scent mark using urine and faeces, and this may be used to mark the home range boundary (Cypher 2003). Gray Foxes have also been observed to use latrine sites, where many animals deposit faeces (Trapp 1978). In Mississippi, Chamberlain and Leopold (2000) found neighbouring males and females did not interact closely (within 150m), suggesting avoidance within this distance, and possibly territoriality.
Published estimates of home range size are predominantly from the southern United States (Fritzell and Haroldson 1982; Cypher 2003). These range from small (e.g. 0.13 - 0.8 km2) in high quality areas (Richards and Hine 1953; Fuller 1978) to large home ranges (e.g. 28 - 77 km2) in areas where food, water and shelter is scarce (Lord 1961; Nicholson 1982), and may vary seasonally and geographically for both males and females. Ranges for both sexes may increase in the fall and winter. During the rearing period, female home range may decline by 80%, and male range may increase (Follman 1973; Nicholson et al. 1985). In Mississippi, 37 Gray Foxes were radio-monitored from 1991 - 1997. Home range did not vary between the sexes, but did vary seasonally (larger during the breeding period 1 Feb - 31 May). During the breeding period home range (95% movements) was 3.5 km2, and 1.8 km2 at other times. Core area size (50% of movements) also varied seasonally, and was largest during the breeding period (0.4 km2), and smallest during the winter period (0.2 km2). Home range overlap was greatest for males and females (57% overlap) and least for adjacent males (1% overlap). Core areas appear to be exclusively used by resident pairs (Chamberlain and Leopold 2000).
Pups forage with their mother at approximately 3 months of age, and independently at 4 months of age (Nicholson et al. 1985). Natal dispersal has been recorded from September to December (Nicholson et al. 1985), with most natal dispersal occurring in autumn (Lord 1961), and likely October (Nicholson et al. 1985). Juveniles remain with the family unit until they are approximately 8 months old (Trapp and Hallberg 1975; Nicholson et al. 1985). The extent of dispersal varies among populations (Cypher 2003). In Alabama, Nicholson et al. (1985) found that 100% of males (N=3) but no females (N=6) dispersed from natal territory, while in New York, 73% of males (N=11) and 63% of females (N=8) dispersed (Tullar and Berchielli 1982).
Dispersal distances of 9, 16, 24 km (Nicholson et al. 1985) and 2 events > 24 km (Tullar and Berchielli 1982) have been recorded for male Gray Fox. The sub-adult male trapped in October in New Brunswick was 135 km north of known records of Gray Fox in Maine (McAlpine et al. 2008).
Males generally disperse further than females; Tullar and Berchielli (1982) recorded average dispersal distances of 7 km (range 0.3 - 22 km) by 8 female Gray Fox, compared to an average of 14 km (range 0.0 - 33 km) by 11 males. Nicholson et al. (1985) only recorded dispersal in males. However, female Gray Fox have also been recorded far from natal territory; Sullivan (1956) recorded a female dispersal event of 83 km, and Sheldon (1953) recovered a marked female 84 km away from her natal area, 3 years after being tagged as a juvenile. One juvenile female in New York moved 18 km in 1 month (Sheldon 1953). Seasonal migration has not been recorded in Gray Fox.
Several predators have been reported to prey on Gray Foxes, including: Golden Eagle, (Aquila chrysaetos) (Mollhagen et al. 1972), Domestic Dog (Tullar and Berchielli 1982), as well as Bobcat (Lynx rufus) and Coyote (Canis latrans) (Fedriani et al. 2000; Farias et al. 2005).
There is evidence that Coyotes have a negative impact on sympatric Gray Fox populations. Coyote and Gray Fox abundance were inversely related in California (Crooks and Soulé 1999; Fedriani et al. 2000), Gray Foxes avoided track stations used by Coyote on the same night (Crooks and Soulé 1999), and Gray Fox numbers increased following Coyote removal in Texas (Henke and Bryant 1999). Coyotes will kill Gray Foxes (Wooding 1984; Farias et al. 2005) and may be a significant predator; 7 of 12 dead radio-tagged Gray Fox in California were killed by Coyote (Fedriani et al. 2000). Habitat and food partitioning, and tree climbing may facilitate some degree of coexistence (Wooding 1984; Cypher 1993) but in areas of high Coyote density Coyote will impact Gray Fox (Fedriani et al. 2000).
There is no evidence that Red Fox outcompete or limit the abundance or distribution of Gray Fox. In Illinois, the amount of overlap in home ranges of Red and Gray Fox overlapped much more than overlap within either species (Follmann 1973). Gray Foxes are more efficient at digesting fruit, and have a narrower gape and smaller carnassials than Red Foxes. These features may allow Gray Foxes to be more effective herbivores, but less effective carnivores, than Red Foxes (Jaslow 1987). Where the two species co-exist, they seem to avoid direct competition because Gray Fox eat mainly plant material and mainly use wooded areas, while Red Fox eat mainly small mammals and use more open, cultivated parts of the same landscape (see Habitat Use). In Maryland, plant consumption by weight was 48% for plants and 35% for mammals in Gray Fox, compared to consumption of 10% plants and 69% mammal in Red Fox (Hockman and Chapman 1983).
Hybridization between Red and Gray Fox seems unlikely because the two species are in separate genera and unlikely to be compatible. There is no evidence of hybridization, except for an unproven record from Ohio, based on a description of a pelt seen 10 years earlier by a fur dealer (Bezdek 1944).
Gray Foxes are resistant to Sarcoptic Mite ('mange', Sarcoptes scabiei) and Heartworm (Dirofilaria immitis), parasites that are fatal or seriously debilitating to most other wild canids, such as Red Fox (Stone et al. 1972; Simmons et al. 1980). The only diseases reported with significant frequency in the Gray Fox are canine distemper and rabies (Gier 1948; Nicholson and Hill 1984; Davidson et al. 1992). Canine distemper and rabies are almost always fatal to Gray Foxes (Fritzell 1987; Yuan Chung Zee 1999) and thus have the potential to limit fox populations under conditions of high Gray Fox density. In a study of 157 sick or dead Gray Foxes from 1972 through 1989, canine distemper was found to be a more significant source of mortality than all other diseases combined (Davidson et al. 1992). In the United States, local population reductions have been observed due to distemper (Nicholson and Hill 1984) and rabies epizootics (Trapp and Hallberg 1975; Steelman et al. 2000).
Evidence of breeding is confined to two sub-populations (Pelee Island, Ontario and Northwestern Ontario). Gray Fox recorded in Alberta, Manitoba, southeastern Ontario, Quebec, and New Brunswick are considered to represent irregular or occasional dispersing animals that are not part of an established breeding population and are not considered significant to the viability of Gray Fox in Canada. Therefore, only records from the two areas of known/probable breeding are used in population estimates.
Population size for all locations is estimated from sightings provided by the public, road-killed individuals, or fur pelt return data. No systematic surveys for Gray Fox have occurred within Manitoba, northwestern Ontario, Quebec, or New Brunswick, and public sightings in these areas are not actively solicited. Gray Fox observations in Manitoba and New Brunswick are not tracked by the provincial Conservation Data Centres because the species is considered to be accidental within these provinces. In Ontario and Quebec, hunting and trapping regulations require reporting of all Gray Fox killed (see Legal Protection and Status). The Ontario Natural Heritage Information Centre records Gray Fox observations in Ontario.
Trail cameras and hair samples collected from scent posts were used to monitor Gray Fox at 30 locations on Pelee Island from 31 May 2012 to 13 May 2013. At each station, one trail camera was established on the forested side of a forest/field edge, and one rubbing station established 5 - 15 m in front of the camera and baited with catnip oil and powdered catnip. Each station was operational for 2 - 4 week periods, multiple times during the year. Hair samples were collected from the rubbing stations (Anon 2012).
Data on population size and trends in the United States are primarily based on hunting and trapping data, although individual trappers are not required to report their catch in any state. Harvest data is often biased because of its dependence on factors such as the number of trappers and hunters, individual effort of trappers, target species (typically species other than Gray Fox, such as Coyote) and market price, and therefore may be only roughly indicative of population trends.
Density estimates for Gray Fox in North America vary between 1.2 - 2.1 animals per km2 (Trapp and Hallberg 1975). However, these estimates are difficult to obtain because foxes have large home ranges, secretive habits, and they are difficult to capture (Cypher 2003). Furthermore, fox density is affected by habitat quality, food availability, interspecific competition, disease and human exploitation. Thus estimation of density may be highly variable spatially (within and among habitats) and temporally (seasonally and annually) (Cypher 2003).
Few data are available on population dynamics in Gray Foxes (Cypher 2003). In Pennsylvania, Richmond (1952) reported a higher number of animals trapped for bounty in years when the period January to March was wetter and warmer than average, and lowest in years when this period was colder and drier; it was assumed that bounty data were correlated with population size.
The frequency of confirmed Gray Fox encounters in Northwestern Ontario appears to have increased during the last 10 years, with sightings most frequently associated with a major road network (Van den Broeck 2014a). After reviewing supporting evidence for Gray Fox observations, roadkill, and trapping records in Northwestern Ontario, Van den Broeck (2014a) estimated that there have been 25 to 28 confirmed Gray Fox individuals reported since 1944, with 21 individuals confirmed in the last 10 years (see Canadian Range). Sightings of a lactating female Gray Fox and multiple foxes travelling together within the last few years implies that breeding in northwestern Ontario is likely. Anecdotal evidence of Gray Fox in southeastern Manitoba indicates that Gray Fox are present, at least in winter, but no data exist to indicate whether there is a resident population of breeding individuals and it is unlikely they are contributing to the Rainy River animals. It is likely that there are < 50 mature animals in the sub-population.
In 2012/2013, Gray Fox were recorded 16 times at eight locations using trail cameras. Seven of these locations were in the northern part of Pelee Island and Gray Fox were most frequently recorded from four of the seven locations (Bowman et al. 2013). The number of individuals is unknown. In 2001, Judge and Haviernick (2002) estimated the Gray Fox population on Pelee Island to be approximately 60 individuals, based on the personal communication of a local hunter and naturalist (N. Beattie) who had seen Gray Fox dens and estimated there were 12 - 15 breeding pairs. If correct, the density would have been 1.4 individuals/km2.
The sub-population is no longer trapped, and protected areas exist where much of the records have been made, suggesting that the earlier estimate of up to 30 mature animals still applies, and may be higher. However, the island is only 42 km2 and Gray Fox have not been reported in greater abundance than previously (Table A3); it is likely that there are < 60 mature animals in the sub-population.
On the adjacent mainland of southwestern Ontario, the last confirmed record was in 2002 (Table A4). It is unlikely that there are more than a few animals, and they likely are individuals dispersing from the United States. This region of Ontario (Essex County) has relatively little forest cover, which may limit Gray Fox persistence in the area.
Sixteen confirmed records of individuals have been reported between 1942 and 1985 along the northeastern shore of Lake Ontario. Two individuals were confirmed between 2000 - 2008, and two individuals in 2014, suggesting Gray Fox may occasionally be dispersing into this area from New York or Quebec (Table A5).
The frequency of confirmed Gray Fox encounters in Quebec appears to have substantially increased in the last 10 years (Dalpé-Charron pers. comm.). There have been 34 confirmed Gray Fox individuals reported since 1894, with 24 individuals confirmed in the last 10 years (see Canadian Range). The number of sightings in the last 5 years and the presence of a male and female trapped in the same area indicates that Gray Fox may be resident in Quebec, although there is no strong evidence of breeding (Table A6). It is likely that there are < 50 mature animals and many individuals are individuals dispersing from the United States.
Two individuals have been reported from New Brunswick (see Canadian Range). These records most likely are of dispersing individuals from Maine. Given the northeastern expansion of the Gray Fox population in Maine (DePue pers. comm.), it is possible that additional dispersing Gray Fox are present, but unreported, from New Brunswick. However, trapping is common in southwestern New Brunswick (Cormier pers. comm. 2014) and if Gray Fox were present in much abundance, there likely would be additional records.
Archeological evidence (Wintemberg 1921, 1928, 1936, 1939, 1948; Downing 1946) suggests that prior to European occupation, the Gray Fox was almost as common as the Red Fox in southern Ontario, as far north as Midland (Downing 1946). Gray Foxes were not recorded in European accounts and it seems Gray Fox were absent from Canada until colonization events of the late 1800s in southern Quebec and late 1940s of southern Ontario (see Canadian Range). A similar extirpation, then recolonization, was recorded in parts of Maine and New England (Waters 1967). The cause of the decline is unknown but corresponded with arrival of Europeans and the related land clearing and unsustainable harvest of many species wild animals (Bozarth et al. 2011). However, Downing (1946) suspected Gray Fox were rare or absent in Ontario at the time of colonization because the species was not mentioned by colonial authors and a large, tree-climbing animal would likely be noticed if it was as abundant as remains from archaeological results suggest.
The extirpation may have been related to climate change; Waters (1964) suggested that Gray Fox range increased north and east during the warm Hypsithermal Period (5000 - 2000 B.C.; Dorf 1959) and then contracted southward as the region cooled. The process repeated during the Medieval Climate Anomaly (800 - 1300 A.D.) when Gray Fox moved into the northeast, before retracting during the Little Ice Age (1500 - 1850) (Bozarth et al. 2011). The mechanism for population change is unknown but Root and Payne (1985) suggested that the smaller average litter size in their northern Wisconsin study may be evidence of less tolerance to adverse environmental and nutritional conditions, and sensitivity to colder environments. The energetic cost of living in areas with deep snow has also been suggested as a limiting factor (Judge and Haviernick 2002).
In the 1930s, Gray Fox expanded northward from the central eastern US states (Hamilton 1943), possibly in association with warming climate and/or increasing amount of young forest on abandoned farmland (Palmer 1956; Bozarth et al. 2011). The arrival of Gray Fox in Northwestern Ontario and Manitoba in the 1940s was thought to be connected to the northward expansion of rabbit (Sylvilagus spp.) (Sutton 1958). The northward expansion of Gray Fox in Wisconsin and Minnesota is thought to be related to climate change (Judge and Haviernick 2002), but may also possibly be related to changes in forest cover associated with periods of intensive timber harvest (Erb pers comm. 2014). In Maine, Gray Fox were absent in the 1940s (Hamilton 1943) but are relatively common in the southern half of Maine today; an average of 134 animals were trapped annually from 2001 - 2006 (McAlpine et al. 2008).
The distribution of Gray Fox in Canada is along the southern border of central-eastern Canada and represents the northern edge of Gray Fox range in North America. Records have been too few in Canada to detect recent trends but it is likely that any population trend in Canada is strongly influenced by the expansion and retraction of populations in adjacent US states.
Gray Foxes have been dispersing into Canada from populations in the United States since the early 20th century (Downing 1946) and probably continue to do so. The presence of the Gray Fox in Canada after a 350-year or less absence is due entirely to immigration from the United States (Downing 1946). It is likely that if the Canadian population were to disappear again, that Gray Fox from the northern US likely would eventually re-populate the Canadian range.
Gray Fox appear to be expanding their range in Minnesota (Erb pers. comm. 2014). Prior to the 1980s they were generally restricted to the hardwood forest portions from southern Minnesota through central Minnesota to the northwestern corner (MNDNR 2014). They are rare in the northwestern counties adjoining Manitoba where agricultural development is very intensive (Erb pers. comm. 2014). However, they have been more frequently observed in the northeastern part of Minnesota adjoining Ontario in the last 10 years, and were first reported from this area in the 1980s (Latimer 2014). The expansion of the Gray Fox population in the northeastern counties of Minnesota likely explains the increase in Gray Fox sightings in Northwestern Ontario in the last 10 years.
Gray Fox are present throughout Michigan and populations are stable (Rollings pers. comm. 2014). Gray Fox are also present throughout Ohio, although at low density in areas near Lake Erie (OHDNR 2014a). The state-wide population trend has fluctuated widely, but overall shows a long-term decline (OHDNR 2014a). This decline is strongly influenced by a high number of sightings/1000 hours in the early 1990s; the population may have stabilized in the last 10 years (Prange pers. comm. 2014). A stable population exists along the New York/Ontario border (Schiavone pers. comm. 2014). Immigration from New York into Ontario was recorded in the Niagara Falls area (see Canadian Range).
It is unknown whether Pelee Island receives animals from Ohio, Michigan, or New York, or how often. Animals could potentially reach Pelee Island on ice during winter. Ohio is the closest potential source for Gray Fox to Pelee Island, located 10 km from intervening islands and 12 km from the mainland, compared to 15 km from the tip of Point Pelee National Park, Ontario. From 1972 to 2011, the area between Ohio and Pelee Island formed > 90% ice cover, every year, except one (NOAA 2015; Wong et al. 2012). Ice typically forms from end-December until end-March, with maximum coverage from mid-January to mid-February (Assell 2004). In 1970, a visiting naturalist was told that hunting blinds he observed along the shoreline at Lighthouse and Fish Points were established each winter to kill incoming Coyote and fox that might impact the Island's annual Ring-necked Pheasant (Phasianus colchicus) hunt (Pratt pers. comm. 2015), suggesting there was likelihood of Gray Fox coming to the island across the ice. However, it is not known if the present Gray Fox sub-population on Pelee Island can be rescued from animals originating from Ohio because Gray Fox populations have been declining in Ohio since the 1990s, and relative abundance is lowest in the areas closest to Canada (Ohio DNR 2015). The frequency of continuous ice between Ohio and Ontario in future years is uncertain.
Movement of Gray Fox from New York to southeastern Ontario was first recorded in 1942 (see Canadian Range) and Gray Fox are presently entering Quebec from New York, Vermont, New Hampshire and Maine. Gray Fox occur throughout Vermont, although they are most numerous in southern Vermont. Trapping data indicate catch-per-unit-effort has increased in the last 10 years, suggesting that the population may have increased, although there is no evidence that they are expanding northward (Bernier pers. comm. 2014). In the Vermont Wildlife Management Units bordering Quebec, a total of 15 Gray Fox have been reported harvested since 1990, with seven of these harvested since 2010 (Bernier pers. comm. 2014).
Based on trapping data, Gray Fox are found throughout New Hampshire, although both trapping effort and capture rates are lowest in northern New Hampshire. There is no evidence of a population increase or range expansion in New Hampshire (Tate pers. comm. 2014).
Gray Fox have been expanding their range in the northeast of Maine for the last 20 years (McAlpine et al. 2008). Their current range extends as far east as the Penobscot River, with some pioneering animals east of this area towards New Brunswick (Figure 2; DePue pers. comm. 2014). It is plausible that sightings of Gray Fox in New Brunswick will increase as the Maine population expands towards New Brunswick. The northern extent of the range in Maine extends to the Madrid area, which is approximately 90 km from the Quebec border.
Range expansion likely is limited by high Coyote densities (see Interspecific Interactions), loss of forest cover (see Habitat Use), and possibly deep snow (see Fluctuations and Trends). However, there is no population-level data on how these factors affect Gray Fox in Canada, or in the US. Much of the loss of forest cover occurred in the 19th century and impacts on recent populations would be difficult to quantify. Milder winters in the future along the border from Manitoba to New Brunswick likely will facilitate Gray Fox expansion into Canada, but deep snow projected for New Brunswick may constitute a problem in that area.
An IUCN Threats Calculator exercise summarized known or predicted threats to Gray Fox in Canada (Appendix 2). Emphasis is given to the two sub-populations where breeding is known, or likely and the threats to these sub-populations are less evident. In other parts of Canada, apparently non-breeding animals are impacted by trapping mortality. It is difficult to apply this scenario to the Threats Calculator; human-caused mortality is not considered to be a limiting factor, but threats are focused on the mature (breeding) cohort. In this report, trapping was considered to be a threat because it may be impacting mature animals, and is likely preventing establishment of more breeding animals. The overall Threat rank was high, due to the impact of trapping mortality in much of the extent of occurrence. The following categories are considered important to the viability of Gray Fox in Canada:
Hunting and Trapping (iucn 5.1)
Records of Gray Fox in Canada are found near the Canada-United States border and it appears that the Gray Fox population in the United States is expanding northward (see Distribution). Few animals have any evidence of breeding and when gender is known, most of the records are of males, suggesting these are dispersing animals emigrating from the United States. Except for records on Pelee Island, many of the Canadian records are obtained from animals killed accidentally by fur harvesters. In Quebec, 24 of 30 (80%) Gray Fox records in the last 20 years were incidentally trapped (MFFP unpub. data; Table A6). Five of 17 Gray Fox observations in northwestern Ontario since 1944 (Van den Broeck 2014a; Table A2), and 1 of 2 New Brunswick records (table A7), were obtained as incidental capture during trapping. Although Gray Fox populations can withstand high harvest pressure (see Life Cycle and Reproduction), incidental capture on the low density populations in Canada likely is limiting the establishment of Gray Fox breeding populations in parts of Canada.
Judge and Haviernick (2002) reported that approximately six to seven Gray Fox were harvested each year from the Whiteshell Provincial Park region in southeastern Manitoba. However, no pelts have been traded recently in Manitoba (Berezanski pers. comm. 2014). Quebec lists Gray Fox as a furbearer so that managers can rapidly introduce regulations if necessary. Quebec's regulations stipulate that live Gray Fox must be released and dead Gray Fox must be reported to a wildlife conservation officer (MFFP 2014).
The most important cause of mortality of Gray Foxes in the United States is human harvest by trapping and hunting (Cypher 2003). Gray Foxes are harvested in all Great Lake and northeastern US states ( Table 1 ). The harvest season ranges from September 1 to March 31 in New Hampshire, and November 3 to December 31 in Maine. There are no restrictions on harvest level. The extent to which harvest levels in the US affect rescue effect for Gray Fox emigrating to Canada is unknown.
A bounty on Gray Fox on Pelee Island existed until the 1980s when it was removed because it did not comply with Ontario's Game and Fish Act (Judge and Haviernick 2002). A resident hunter and naturalist, N. Beattie, is cited in the 2002 COSEWIC report stating that hunters on Pelee Island shot six to ten Gray Foxes every winter in the 1990s, and that there was no indication that this practice had a negative effect on the population (Judge and Haviernick 2002). There is no recent information on the existence, or extent, of illegal or accidental harvest of Gray Fox on Pelee Island.
| State | Hunting season | Trapping season | Bag limit | Source |
|---|---|---|---|---|
| Wisconsin | Oct 19-Feb 14 | Oct 19-Feb 14 | No limit | WIDNR 2013 |
| Michigan | Oct 15-Mar 1 | Oct 15-Mar 1 | No limit | MIDNR 2013 |
| Ohio | Nov 10-Jan 31 | Nov 10-Jan 31 | No limit | OHDNR 2014b |
| New York | Oct 25-Feb 15 | Oct 25-Dec 10 Table Footnotea Oct 25-Feb 15 Table Footnoteb |
No limit | NYDEC 2014 |
| Vermont | Oct 25-Feb 8 | Oct 25-Dec 31 | No limit | VTDFW 2013 |
| New Hampshire | Sep 1-Mar 31 | Oct 15-Mar 1 | No limit | NHFGD 2014 |
| Maine | Oct 20-Feb 28 | Nov 3-Dec 31 | No limit | MEDIFW 2013 |
Residential and Commercial Development (iucn 1.0)
A study in southern California found Gray Fox were negatively associated with increased urbanization, and proximity to urban areas (Ordenana et al. 2010.) However, the study was conducted in an area of high human density (19 million people) and it is unlikely that the effect of urbanization would be realized within Canadian range of Gray Fox.
Roads (iucn 4.1)
Roadkill is considered a limiting factor; its status as a threat is unclear.
Gray Foxes are susceptible to road mortality due to their large home range, large dispersal distances, and association with rural landscapes. Data on road mortality in Canada do not exist. However, one study in Louisiana that radio-tracked Gray Foxes found 8 of 17 deceased foxes were killed in vehicle collisions. They estimated that Gray Foxes had a 34% probability of experiencing human-induced mortality (road mortality and trapping combined).
In Quebec, 3 of 34 Gray Fox observations were obtained as roadkill (MFFP unpub. data). At least three Gray Fox observations were obtained as roadkill in eastern Ontario. There are anecdotal reports of Gray Fox killed on roads in southeastern Manitoba (see Distribution).
In the two sub-populations with breeding, 3 of 25 Gray Fox observations were obtained as roadkill in the Northwestern Ontario sub-population, and 2 of 17 observation events in the Pelee Island sub-population. Traffic volume on Pelee Island is relatively low, but roadkill does constitute a source of mortality.
Invasive Non-native Disease (iucn 8.1)
Non-native diseases such as canine distemper and rabies are fatal to Gray Foxes and have been reported in significant numbers of wild individuals in the United States (Davidson et al. 1992). In an epizootic situation these diseases could represent significant limiting factors. In Ontario, two Gray Fox had rabies; both cases were in 1986, a peak year, with the highest number of rabies cases of all species in Ontario to date (Bachmann pers. comm. 2014). The prevalence of any disease in Canadian Gray Foxes is unknown because few animals have been assessed.
The number of locations is unknown but likely exceeds 10, which is the threshold for using location number in a status assessment. The breeding population on Pelee Island is contained within a small (42 km2) area and threats are similar across this sub-population, suggesting the sub-population is a single location. However, the Northwestern Ontario sub-population resides in a larger area (approximately 35,000 km2) and trapping effort likely varies enough that numerous locations exists. It is noted that the concept of location is difficult to apply to a vagile species whose distribution is based on rare, scattered, point-source records; associating these records with a geographic or ecologically distinct area is problematic in identifying a specific threat.
The Gray Fox was assessed by COSEWIC as Threatened in 2015 (and prior to that, Threatened in 2002). This species is currently listed as Threatened on Schedule 1 of the federal Species at Risk Act (SARA). The Canada National Parks Act protects Gray Fox and their habitat within National Parks (e.g. Point Pelee National Park, St Lawrence Islands National Park, and Trent-Severn Waterway Historic Site).
In Ontario, the Gray Fox was listed as Threatened under the Ontario Endangered Species Act in 2008 and the species and its habitat are protected. In Quebec, the Gray Fox is not listed as threatened, vulnerable, or likely to be threatened or vulnerable in the Act Respecting Threatened or Vulnerable Species. In both Ontario and Quebec, hunting and trapping guidelines require Gray Fox that are incidentally trapped to be released if alive, or reported (and potentially surrendered) to conservation officers if deceased (MRNF 2014a,b, Ontario Fish and Wildlife Conservation Act). Ontario's Forest Management Guidelines for Conserving Biodiversity at the Stand and Site Scales (OMNR 2010) provides protection for occupied Gray Fox den sites known before, or found during, forest management operations.
In Manitoba and New Brunswick, records of Gray Fox are considered to be accidental and not protected by the Manitoba Endangered Species Act, 2007, or the New Brunswick Species at Risk Act. They are not considered a furbearer under the Manitoba Wildlife Act. In New Brunswick, regulations for harvest of Gray Fox would be the same as for Red Fox because Gray Fox were not considered to be part of New Brunswick, and the Fish and Wildlife Act refers only to "fox".
Globally, the Gray Fox is ranked as Least Concern by the IUCN because it is widespread and there is no evidence that their numbers are decreasing in any part of their range (Cypher et al. 2008). NatureServe (2014) ranks the species as Secure globally (G5) and in the United States (N5). In the states adjacent to Canadian occurrences, the Gray Fox is ranked as Secure (S5: Maine, Vermont, New York, Pennsylvania), Apparently Secure to Secure (S4S5: New Hampshire, Wisconsin), Apparently Secure (S4: Michigan) or unranked (SNR: Ohio, Minnesota). Other state ranks are listed in Table 2 .
In Canada and Ontario, NatureServe ranks the Gray Fox as Critically Imperiled (N1, S1) and the Wild Species Report ranks the species as At Risk (Rank 1, CESCC 2006, 2011). It is considered accidental in Alberta, Manitoba, and New Brunswick (CESCC 2011), and is therefore unranked by both NatureServe and the Wild Species Report. In Quebec, the Gray Fox is considered 'occasional' with a NatureServe rank of SNA (Gauthier pers. comm. 2014; MFFP 2014c).
| NatureServe rank | Description | US state |
|---|---|---|
| S2 | Imperiled | Wyoming |
| S3 | Vulnerable | District of Columbia, Iowa, Kansas |
| S3S4 | Vulnerable-Apparently secure | Utah |
| S4 | Apparently secure | Colorado, Indiana, Kentucky, Michigan, Missouri, Nebraska, Oklahoma, Oregon |
| S4S5 | - | Louisiana, New Hampshire, Wisconsin |
| S5 | Secure | Alabama, Arizona, Arkansas, Connecticut, Delaware, Georgia, Illinois, Maine, Maryland, Massachusetts, Mississippi, Navajo Nation, Nevada, New Jersey, New Mexico, New York, North Carolina, Pennsylvania, Rhode Island, South Dakota, Tennessee, Texas, Vermont, Virginia, West Virginia |
| SU | Under review | North Dakota |
| SNR | Not ranked | California, Florida, Minnesota, Ohio, South Carolina |
Gray Fox records in Manitoba and northwestern Ontario are primarily found on private land or crown land managed for timber production. In Manitoba, Gray Fox have been reported from Whiteshell Provincial Park. No specific habitat protection measures are established for Gray Fox.
Gray Fox habitat on Pelee Island occurs in Fish Point Provincial Nature Reserve, managed by Ontario Parks under the Provincial Parks and Conservation Reserves Act and Stone Road Alvar managed by the Essex Region Conservation Authority under the Conservation Authority Act. Gray Fox habitat is also protected on over 400ha of privately owned land managed for conservation purposes by Ontario Nature and the Nature Conservancy of Canada.
Gray Fox records are primarily found on private land or crown land. There are (unconfirmed) historical reports of Gray Fox in St. Lawrence Islands National Park, Ontario and a single animal was killed there in 2008 (Parks Canada 2014), Point Pelee National Park, and the Trent-Severn Waterway National Historic site, but no recent reports from these locations (Nantel pers. comm. 2014).
Dean Berezanski, Provincial Furbearer Biologist, Manitoba Conservation and Water Stewardship, Winnipeg, Manitoba
Chris Bernier, Furbearer Biologist, Vermont Fish and Wildlife, Springfield, Vermont.
Sean Blaney, Botanist and Executive Director, Atlantic Canada Conservation Data Centre, Sackville New Brunswick
Jeff Bowman, Research Scientist, Wildlife Research and Monitoring Section, Ontario Ministry Natural Resources and Forestry, Peterborough, Ontario.
Jennifer Boyko, Management Biologist, Dryden District, Ontario Ministry Natural Resources, Dryden
Alain Branchaud. Species at Risk Recovery Biologist. Environment Canada, Montréal, Quebec.
Vivian Brownell, Senior Species at Risk Biologist, Species at Risk Branch, Ontario Ministry Natural Resources, Peterborough, Ontario
Emmanuel Dalpé-Charron, Biologiste et coordonnateur provincial, Ministère des Forêts, de la Faune et des Parcs du Québec, Québec, Québec.
Michael Deschamps, Area Biologist, Thunder Bay South Area, Ontario Ministry of Natural Resources, Thunder Bay, Ontario.
John DePue, Furbearer/Small Mammal Biologist. Maine Inland Fisheries and Wildlife, Bangor, Maine.
Dave Duncan, Manager, Population Conservation Section, Canadian Wildlife Service, Environment Canada, Edmonton, Alberta.
John Erb, Furbearer Research Biologist, Minnesota Department of Natural Resources, Grand Rapids, Minnesota.
Chris Friesen, Biodiversity Information Manager, Manitoba Conservation Data Centre, Winnipeg, Manitoba.
Martina Furrer, Biodiversity Information Biologist, Natural Heritage Information Centre, Ontario Ministry of Natural resources, Peterborough, Ontario.
Isabelle Gauthier, Coordonnatrice provinciale, espèces fauniques menacées et vulnérables, Ministère des Forêts, de la Faune et des Parcs du Québec, Québec.
Neil Jones, Scientific Project Officer and ATK Coordinator, COSEWIC secretariat, Canadian Wildlife Service, Environment Canada, Gatineau Québec
Kimberley McNaughton, Management Biologist, Nipigon District, Ontario Ministry Natural Resources
Don McAlpine, Head, Department of Natural Science, New Brunswick Museum, Saint John, New Brunswick.
Christy McDonald, Wildlife Policy Analyst, Wildlife Policy Section, Ontario Ministry Natural Resources, Peterborough
Mhairi McFarlane, Conservation Biologist, southwestern Ontario, The Nature Conservancy of Canada, London, Ontario.
Randall Mooi, Curator of Zoology, The Manitoba Museum, Winnipeg, Manitoba.
Patrick Nantel, Office of the Chief Ecosystem Scientist, Parks Canada, Gatineau, Quebec
Michael Oldham, Ontario Natural Heritage Information Centre (NHIC), Ontario Ministry of Natural Resources, Peterborough, Ontario.
Suzanne Prange, Furbearer Biologist, Ohio Department of Natural Resources, Ohio.
Brian Rollings, Wildlife Biologist, Michigan Department of Natural Resources, Marquette, Michigan.
Rich Russell, Canadian Wildlife Service, Environment Canada, Downsview, Ontario.
Michael Schiavone, Certified Wildlife Biologist, New York State Department of Environmental Conservation, Albany, New York.
Irene Schneider, Ontario Ministry of Natural Resources, Dryden, Ontario.
Sonia Schnobb, Administrative Assistant, COSEWIC Secretariat, Canadian Wildlife Service, Environment Canada, Gatineau, Québec.
Jessica Sicoly, Management Biologist, Ontario Ministry of Natural Resources and Forestry, Sault Ste Marie, Ontario.
Don Sutherland, Ontario Natural Heritage Information Centre (NHIC), Ontario Ministry of Natural Resources, Peterborough, Ontario.
Patrick Tate, Furbearer Project Leader, New Hampshire Fish and Game, Concord, New Hampshire.
John Van den Broeck, Biodiversity Species at Risk Biologist, Ontario Ministry of Natural Resources, Fort Frances, Ontario.
Bill Watkins, Biodiversity Conservation Zoologist, Wildlife Branch, Manitoba Conservation and Water Stewardship, Winnipeg, Manitoba.
Jenny Wu, Scientific Project Officer, COSEWIC Secretariat, Canadian Wildlife Service, Environment Canada, Gatineau, Québec.
Anderson, R.M. 1939. Mammifères de la province de Québec (revisée). The Provencher Society of Natural History of Canada, 1939 Annual Report. pp.37-111.
Anon. 2012. Scent camera station protocol - Pelee Island Grey Fox project, 8 May 2012. Nature Conservancy of Canada.
Arscott, C. pers. comm. 2009. Email correspondence to Environment Canada (SARA registry website). September 1, 2009. Catchacoma, Ontario.
Ashton, K.G., M.C. Tracy, and A. deQueroz. 2000. Is Bergmann's rule valid for mammals? American Naturalist 156: 390-415.
Assell, R. 2004. Lake Erie Ice Cover Climatology 1898-2002. NOAA Technical Memorandum GLERL - 128. [Accessed August 2015].
Aubry, K. B., M.J. Statham, B.N. Sacks, J.D. Perrine, and S.M. Wisely. 2009. Phylogeography of the North American red fox: Vicariance in Pleistocene forest refugia. Molecular Ecology 18: 2668-2686.
Bachmann, pers. comm. 2014. Email correspondence to J. Bowman. November 2014. Senior Wildlife Research Technician, Ontario Ministry of Natural Resources and Forestry, Peterborough, Ontario.
Bachmann, P., and W. Lintack. 1982. Movement and activity of a grey fox (Urocyon cinereoargenteus) in Ontario. Ontario Ministry of Natural Resources Rabies Research Unit Unpublished Report. 5 pp.
Banfield, A.W.F. 1974. The Mammals of Canada. University of Toronto Press, Toronto. xxv + 437 pp.
Bangs, O. 1899. A new Gray Fox from the upper Mississippi valley. Proceedings of the New England Zoological Club 1: 43-33.
Berezanski, D. pers. comm. 2014. Email correspondence to J. Pearce. February 2014. Provincial Furbearer Biologist, Manitoba Conservation and Water Stewardship, Winnipeg, Manitoba.
Bernier, C., pers. comm. 2014. Telephone conversation with J. Pearce. July 2014. Furbearer Biologist, Vermont Fish and Wildlife, Springfield, Vermont.
Bezdek, H. 1944. A red-grey fox hybrid. Journal of Mammalogy 25:90.
Bowman, J., pers. comm. 2015. Written comments on draft report to Jennie Pearce, April 2015. Ontario Ministry of Natural Resources, Peterborough, Ontario.
Bowman, J., T.L. Pulfer, L. Bruce, M. McFarlane, C. Sadowski, J. Croswaithe and J. Nageleisen. 2013. Gray Fox (Urocyon cinereoargenteus) monitoring in Pelee Island. Unpublished report. Ontario Ministry of Natural Resources, Peterborough, Ontario. 2pp.
Bozarth, C.A., S.L. Lance, D.J. Civitello, J.L. Glenn, and J.E. Maldonado. 2011. Phylogeography of the gray fox (Urocyon cinereoargenteus) in the eastern United States. Journal of Mammalogy 92: 283-294.
Bruce, L., pers. comm. 2015. Results of Grey Fox necropsies reported to Jennie Pearce. Ministry of Natural Resources and Forestry, Peterborough, Ontario.
Campbell, C.A., A.I. Dagg, M. Dyer, and M.E. Gartshore. 1972. Mammals of Waterloo and South Wellington Counties 1972. Privately Printed, Waterloo, ON. 130 pp.
Canadian Endangered Species Conservation Council (CESCC). 2006.
Wild Species 2005: The General Status of Species in Canada.
Canadian Endangered Species Conservation Council (CESCC). 2011. Wild Species 2010: The General Status of Species in Canada. National General Status Working Group: 302 pp. [Accessed 17 June 2014].
Chamberlin, M.J., and B.D. Leopold. 2000. Spatial use patterns, seasonal habitat selection, and interactions among adult gray foxes in Mississippi. Journal of Wildlife Management 64: 742-751.
Cormier, J. pers. comm. (2014). Email correspondence to J. Pearce. November 2014. Furbearer Management Biologist, Department of Natural Resources, Fredericton, New Brunswick.
COSEWIC. 2002. COSEWIC assessment and update status report on the grey fox Urocyon cinereoargenteus interior in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. vi + 32 pp.
Crooks, K.R. and M.E. Soulé. 1999. Mesopredator release and avifaunal extinctions in a fragmented system. Nature 400: 563-566.
Cypher, B.L. 1993. Food item use by three sympatric canids in southern Illinois. Transactions of the Illinois State Academy of Science 86: 139-144.
Cypher, B.L. 2003. Foxes (Vulpes species, Urocyon species, and Alopex lagopus). Pp 511-546. in G.A. Feldhamer, B.C. Thompson, and J.A. Chapman (eds.). Wild Mammals of North America: Biology, Management and Conservation. John Hopkins University Press, Baltimore, Maryland.
Cypher, B.L., T.K. Fuller, and R. List. 2008. Urocyon cinereoargenteus in IUCN 2013. IUCN Red List of Threatened Species. Version 2013.2. [Accessed 26 April 2014].
Dalpé-Charron, E. pers. comm. 2014. Email correspondence to J. Pearce. Biologiste et coordonnateur provincial, Ministère des Forêts, de la Faune et des Parcs du Québec, Québec, Québec.
Davidson, W.R., V.F. Nettles, L.E. Hayes, E.W. Howerth, and C.E. Couvillion. 1992. Diseases diagnosed in gray foxes (Urocyon cinereoargenteus) from the southeastern United States. Journal of Wildlife Diseases 28: 28-33.
Davis, W.B. 1960. The Mammals of Texas (Bulletin 41). Texas Game and Fish Commission, Austin, Texas.
Davis, D.E., and J.E. Wood. 1959. Ecology of foxes and rabies control. Public Health Report 74: 115-118.
DePue, J. pers.. comm. 2014. Email correspondence to J. Pearce. May 2014. Furbearer/Small Mammal Biologist, Maine Inland Fisheries and Wildlife, Bangor, Maine.
Deschamps, Michael. Pers. comm. September 2015. Email correspondence with Graham Forbes. Area Biologist, Ontario Ministry of Natural Resources, Thunder Bay.
Dorf, E. 1959. Climatic changes of the past and present. Contributions from Museum of Paleontology, University of Michigan 8:181-210.
Downing, S.C. 1946. The history of the grey fox in Ontario. Canadian Field-Naturalist 60: 45-46.
Ennen, J., M.E. Kalis, A.L. Patterson, B.R. Kreiser, J.E. Lovich, J. Godwin, and C.P. Qualls. 2014. Clinal variation or validation of a subspecies? A case study of the Graptemys nigrinoda complex. Biological Journal of the Linnean Society. 111: 810-822.
Erb, J. pers comm. 2014. Telephone conversation with J. Pearce. May 2014. Furbearer Research Biologist, Minnesota Department of Natural Resources, Grand Rapids, Minnesota.
Essex Region Conservation Authority 2011. Watershed characterization. Updated/Amended Proposed Assessment Report. Essex Region Conservation Authority. Website: http://www.essexregionsourcewater.org/downloads/ar_chapter2.pdf. [Accessed:July 2014].
Farias, V., T.K. Fuller, R.K. Wayne, and R.M. Sauvajot. 2005. Survival and cause-specific mortality of gray foxes (Urocyon cinereoargenteus) in southern California. Journal of Zoology 266: 249-254.
Fedriani, J. M., T. K. Fuller, R. M. Sauvajot, and E. York. 2000. Competition and intraguild predation among three sympatric carnivores. Oecologia 125: 258-270.
Follmann, E.H. 1973. Comparative ecology and behavior of red and gray foxes. Ph.D. Thesis. University of Carbondale, Southern Illinois, IL.
Follmann, E.H. 1978. Annual reproductive cycle of the male gray fox. Transactions of the Illinois State Academy of Science 71: 304-311.
Fritzell, E.K. 1987. Gray fox and island gray fox. pp. 409-420 in M. Novak, J.A. Baker, M.E. Obbard, and B. Malloch (eds.). Wild Furbearer Management and Conservation in North America. Ontario Trappers Association, North Bay, ON. viii + 1150 pp.
Fritzell, E.K., and K.J. Haroldson. 1982. Urocyon cinereoargenteus. Mammalian Species 189: 1-8.
Fuller, T. 1978. Variable home rang sizes of female Gray Foxes. Journal of Mammalogy 59:447-448.
Gauthier, I. pers. comm. 2014. Email correspondence with G. Forbes. March 2015. Biologist, Coordonnatrice provinciale,espèces fauniques menaces et vulnérables, Ministère des Forêts, de la Faune et des Parcs, Québec City, Québec.
Gier, H.T. 1948. Rabies in the wild. Journal of Wildlife Management 12: 142-153.
Grinnell, J., J.S. Dixon, and J.M. Linsdale. 1937. Fur-bearing Mammals of California, Volume II. University of California Press, Berkeley, CA. xiv + 777 pp.
Hallberg, D.L., and G.R. Trapp. 1984. Gray fox temporal and spatial activity in a riparian-agricultural zone in California's Central Valley. pp. 920-928 in R.W. Warner and K.M. Hendrix (eds.). Proceedings of the California riparian systems conference. University of California Press, Berkeley California.
Hall, E.R. 1981. The Mammals of North America, Volume II. John Wiley & Sons, New York, NY. xv + 1181 + 90 pp.
Hall, E.R. 1984. Geographic variation among brown and grizzly bears (Ursus arctos) in North America. University of Kansas Special Publication 13. Lawrence, Kansas.
Hamilton, W.J., Jr. 1943. The Mammals of Eastern United States. Comstock Publishing Co., Ithaca, NY.
Haroldson, K.J., and E.K. Fritzell. 1984. Homes ranges, activity, and habitat use by gray foxes in an oak-hickory forest. Journal of Wildlife Management 48: 222-227.
Hatfield, D.M. 1939. Winter food habits of foxes in Minnesota. Journal of Mammalogy 20: 202-206.
Henke, S.E., and F.C. Bryant. 1999. Effects of coyote removal on the faunal community in western Texas. Journal of Wildlife Management 63: 1066-1081.
Hockman, G., and J. Chapman. 1983. Comparative feeding habits of red foxes (Vulpes vulpes) and gray foxes (Urocyon cinereoargenteus) in Maryland. American Midland Naturalist 110:276-285.
IUCN 2010. Guidelines for application of IUCN Red List Criteria at regional and national levels. Version 4.0. Prepared by the IUCN Species Survival Commission, January 2010. PDF.
IUCN 2014. Guidelines for Using the IUCN Red List Categories and Criteria. Version 11. Prepared by the Standards and Petitions Subcommittee. http://www.iucnredlist.org/documents/RedListGuidelines.pdf.
Jackson, H.H.T. 1961. Mammals of Wisconsin. University of Wisconsin Press, Madison, WI. xii + 504 pp.
James, F. 2010. Avian subspecies: Introduction. Ornithological Monographs 67:1-5.
Jaslow, C.R. 1987. Morphology and digestive efficiency of red foxes (Vulpes vulpes) and grey foxes (Urocyon cinereoargenteus) in relation to diet. Canadian Journal of Zoology 65: 72-79.
Jolicoeur, H., P-Y. Collin, N. Latour and P. Canac-Parquis. 2011. Mise à jour des mentions de renard gris (Urocyon cinereargenteus) dans le sud du Québec. Le Naturaliste Canadien 135: 74-81.
Judge, K., and M. Haviernick. 2002. Update status report on the grey fox Urocyon cinereoargenteus in Canada. COSEWIC 32 pp.
Kutschera, V., N. Lecomte, A. Janke, N. Selva, A.A. Sokolov, T. Haun, K. Steyer, C. Nowak, and F. Haller. 2013. A range-wide synthesis and timeline for phylogeographic events in the red fox (Vulpes vulpes). BMC Evolutionary Biology 13:114.
Latham, R.M. 1943. An ecological study of the red and gray foxes in southeastern Pennsylvania (Pittman-Robertson Report I-R). Pennsylvania Game Commission, Harrisburg, Pennsylvania.
Latimer, J. 2014. Northern Minnesota phenology for February 2014: Gray Fox. [Accessed June 2014]
Layne, J.N. 1958. Reproductive characteristics of the gray fox in southern Illinois. Journal of Wildlife Management 22: 157-163.
Layne, J.N., and W.H. McKeon. 1956a. Notes on red and gray fox den sites in New York. New York Fish and Game Journal 3: 248-249.
Layne, J.N., and W.H. McKeon. 1956b. Some aspects of red fox and gray fox reproduction in New York. New York Fish and Game Journal 3: 44-74.
Lindbald-Toh, K., (and 46 co-authors). 2005. Genome sequence, comparative analysis and haplotype structure of the domestic dog. Nature 438:803-819.
Linnitt, A. pers. comm. 2009. Email correspondence to Environment Canada (general inquiries address). June 1, 2009. Minto, Ontario.
London Free Press. 1998. Rare grey fox discovered. London Free Press, Monday, 15 June 1998.
Lord, R.D., Jr. 1961. A population study of the gray fox. American Midland Naturalist 66: 87-109.
MacDonald, I.D. 1989. A Biological Inventory and Evaluation of the Oriskany Sandstone and Woodlands Area of Natural and Scientific Interest. Parks and Recreational Areas Section, OMNR, OFER 8903, Central Region, Richmond Hill, ON. x + 193 pp.
Maine Department of Inland Fisheries and Wildlife (MEDIFW) 2013. Maine Department if Inland Fisheries and Wildlife. [Accessed 1 July 2014].
Manitoba Wildlife Branch. 2015. Fur bearing animal harvest and values. Wildlife Branch. Government of Manitoba. . [Accessed August 2015].
McAlpine, D. pers. comm. 2014. Email correspondence to J. Pearce. June 2014. Head, Department of Natural Science, Research Curator and Head, Zoology Section, New Brunswick Museum, Saint John, New Brunswick.
McAlpine, D., J.D. Martin, and C. Libby. 2008. First occurrence of the grey fox, Urocyon cinereoargenteus, in New Brunswick: a climate-change mediated range expansion? Canadian Field-Naturalist 122: 169-171.
McAlpine, D.F, J. Cormier, and I.G. McLean. (2016). Second report of the Grey Fox, Urocyon cinereoargenteus, in New Brunswick and its conservation significance. Canadian Field-Naturalist.
McFarlane, M., unpub. data. 2013. Unpublished data. The Nature Conservancy of Canada, London, Ontario.
McFarlane, M. 2013. What does the fox say? Well that really wasn't the focus of the study. Nature Conservancy of Canada, Toronto, Ontario. [Accessed 21 April 2015].
Merriam, C.H. 1903. Eight new mammals from the United States. Proceedings of the Biological Society of Washington 16: 73-78.
Merriam, C.H. 1918. Review of the Grizzly and Big Brown Bears of North America. North America Fauna 41:1-136.
Michigan Department of Natural Resources (MIDNR). 2013. 2013 Hunting and Trapping Digest. Michigan Department of Natural Resources, Lancing, MI. [Accessed 1 July 2014].
Minnesota Department of Natural Resources (MNDNR) 2014. Mammals of Minnesota: Grey Fox. Minnesota Department of Natural Resources, St. Paul, MN. [Accessed 15 May 2014].
Ministères des Forêts, de la Faune et des Parcs du Québec (MFFP). 2014. Unpublished data. Ministeres des Forets, de la Faune et des Parcs du Québec, Québec, Québec.
Ministères des Forêts, de la Faune et des Parcs du Québec (MFFP). 2014a. Trapping in Québec, 2014-2016. [Accessed 21 April 2015].
Ministères des Forêts, de la Faune et des Parcs du Québec (MFFP). 2014b. Sport Hunting in Québec 2014-2016. Ministère des Ressources naturelles et de la Faune, Québec City, Québec. Web site: http://mffp.gouv.qc.ca/english/publications/online/wildlife/hunting-regulations/index.asp. [Accessed 21 April 2015].
Ministères des Forêts, de la Faune et des Parcs du Québec (MFFP). 2014c. Liste de la faune vertébrée du Québec. [French only, Accessed 22 April 2014].
Mollhagen, T.R., R.W. Wiley, and R.L. Packard. 1972. Prey remains in golden eagle nests: Texas and New Mexico. Journal of Wildlife Management 36: 784-792.
Moore, J.E. 1952. The gray fox in Alberta. Journal of Mammalogy 33: 253.
Nantel, P. pers. comm. 2014. Email correspondence to J. Pearce. February 2014. Office of the Chief Ecosystem Scientist, Parks Canada, Gatineau, Québec.
NatureServe. 2014. NatureServe Explorer: An online encyclopedia of life [web application]. Version 7.1. NatureServe, Arlington, Virginia. Web site: http://www.natureserve.org/explorer. [Accessed February 2014].
New Hampshire Fish and Game Department (NHFGD). 2014. 2013 - 2014 New Hampshire Hunting and Trapping Digest. New Hampshire Fish and Game Department, Concord, NH. [Accessed 1 July 2014].
Niagara Region Conservation Authority. 2013. Updated Assessment Report: Niagara Peninsula Source Protection Area. Niagara Peninsula Conservation Authority, Welland, ON. [Accessed: July 2014].
Nicholson, W. S. 1982. An ecological study of the gray fox in east-central Alabama. M.S. Thesis Auburn University, Auburn, Alabama. 93pp.
Nicholson, W.S. and E.P. Hill. 1984. Mortality in gray foxes from east-central Alabama. Journal of Wildlife Management 448: 1429-32.
Nicholson, W.S., E.P. Hill, and D. Briggs. 1985. Denning, pup-rearing, and dispersal in the gray fox in east-central Alabama. Journal of Wildlife Management 49: 33-37.
Nowak, R.M. 1995. Another look at wolf taxonomy. Pp 375-397 in L. Carbyn, S. Fritts, and D. Seip (eds.). Ecology and Conservation of Wolves in a Changing World. Canadian Circumpolar Institute, Alberta. 620 pp.
New York Department of Environment and Conservation (NYSDEC). 2014. Hunting Seasons. New York State Department of Environmental Conservation [Accessed 1 July 2014].
NOAA 2015. Lake Erie Ice Cover [Accessed August 2015].
Ohio Department of Natural Resources (OHDNR). 2014a. Gray Fox - Urocyon cinereoargenteus. Ohio Department of Natural Resources Division of Wildlife. Website: http://wildlife.ohiodnr.gov/species-and-habitats/species-guide-index/mammals/gray-fox. [Accessed August 2015].
Ohio Department of Natural Resources (OHDNR). 2014b. 2014 - 2015 Season Dates & Bag Limits. Ohio Department of Natural Resources Division of Wildlife. [Accessed 1 July 2014].
Ontario Ministry of Natural Resources 2010. Forest Management Guidelines for Conserving Biodiversity at Stand and Site Scales. Queen's Printer, Toronto, Ontario, 211 pp.
Ontario Partners in Flight. 2008a. Ontario Landbird Conservation Plan: Boreal Hardwood Transition, North American Bird Conservation Region 12. Ontario Ministry of Natural Resources, Bird Studies Canada, Environment Canada. PDF. [Accessed 30 June 2014].
Ontario Partners in Flight. 2008b. Ontario Landbird Conservation Plan: Lower Great Lakes/St. Lawrence Plain, North American Bird Conservation Region 13. Ontario Ministry of Natural Resources, Bird Studies Canada, Environment Canada. PDF. [Accessed 30 June 2014].
Ordenana, M., (and 11 co-authors). Effects of urbanization on carnivore species distribution and richness. Journal of Mammalogy 91:1322-1331.
Palmer, R. S. 1956. Gray fox in the Northeast. Maine Field Naturalist 12:62-70.
Parks Canada. 2014. Managed Area Element Status Assessment. Unpublished report. Parks Canada Agency, Ottawa, Ontario.
Petersen, L.R., M.A. Martin, and C.M. Pils. 1977. Status of gray foxes in Wisconsin, 1975. Wisconsin Department of Natural Resources, Research Report No. 94, Madison, WI. 17 pp.
Peterson, R.L., R.O. Stanfield, E.H. McEwen, and A.C. Brooks. 1953. Early records of the red and the gray fox in Ontario. Journal of Mammalogy 34: 126-127.
Pils, C.M. and W.D. Klimstra. 1975. Late fall foods of the gray fox in southern Illinois. Transactions of the Illinois State Academy of Science 68: 255-262.
Prange, S. pers. comm. 2014. Telephone conversation with J. Pearce. 18 November 2014. Furbearer Biologist, Ohio Department of Natural Resources, Ohio.
Pratt, Paul. pers. comm. 2015. Email correspondence with Graham Forbes, July 2015.
Rausch, R. 1963. Geographic variation in size of North American Brown Bear (Ursus arctos) as indicated by condylobasal length. Canadian Journal of Zoology 41:33-45.
Richmond, N.D. 1952. Fluctuations in gray fox populations in Pennsylvania and their relationship to precipitation. Journal of Wildlife Management 16: 198-208.
Richards, S.W., and R.L. Hines. 1953. Wisconsin fox populations. Wisconsin Conservation Department, Technical Wildlife Bulletin No. 6, Madison, WI. 78 pp.
Rollings, B., pers. comm. 2014. Phone conversation with J. Pearce. July 2014. Wildlife Biologist, Michigan Department of Natural Resources, Marquette, Michigan.
Root, D.A. 1981. Productivity and mortality of gray foxes and raccoons in southwestern Wisconsin. MS Thesis, University of Wisconsin, Stevens Point, Wisconsin.
Root, D.A., and N.F. Payne. 1985. Age-specific reproduction of gray foxes in Wisconsin. Journal of Wildlife Management 49: 890-892.
Sadowski, C., pers. comm. 2015. Results of Grey Fox necropsies reported to Jennie Pearce. Ministry of Natural Resources and Forestry, Peterborough, Ontario.
Schiavone, M., pers comm. 2014. Email and telephone correspondence with J. Pearce. May 2014. Certified Wildlife Biologist, New York State Department of Environmental Conservation, Albany, New York.
Schmeltz, L.L. and J.O. Whitaker, Jr. 1977. Use of woodchuck burrows by woodchucks and other mammals. Transactions of the Kentucky Academy of Science 38: 79-82.
Schreber, J. 1775. Die Säugthiere in Abbildungen nach der Natur mit Beschreibungen. 7 volumes; Plate 92 (Vol. 2); Text 361 (Vol. 3).
Sheldon, W.G. 1949. Reproductive behavior of foxes in New York State. Journal of Mammalogy 30: 236-246.
Sheldon, W.G. 1953. Returns on banded red and gray foxes on New York state. Journal of Mammalogy 34, 125.
Sicoly, J., pers comm. 2014. Phone conversation with J. Pearce. September 2014. Management Biologist, Ontario Ministry of Natural Resources and Forestry, Sault Ste Marie, Ontario.
Simmons, J.M., W.S. Nicholson, E.P. Hill, and D.B. Briggs. 1980. Occurrence of (Dirofilaria immitis) in gray fox (Urocyon cinereoargenteus) in Alabama and Georgia. Journal of Wildlife Diseases 16: 225-228.
Smith, H.C. 1993. Alberta mammals: an atlas and guide. Provincial Museum of Alberta, Edmonton, AB. xii + 239 pp.
Steelman, H.G., S.E. Henke, and G.M. Moore. 2000. Bait delivery for oral rabies vaccine to gray foxes. Journal of Wildlife Diseases 36: 744-51.
Stone, W.B., Jr., E. Parks, B.L. Weber, and F.J. Parks. 1972. Experimental transfer of sarcoptic mange from red foxes and wild canids to captive wildlife and domestic animals. New York Fish and Game Journal 19: 1-11.
Sullivan, E.G. 1956. Gray fox reproduction, denning, range, and weights in Alabama. Journal of Mammalogy 37: 346-51.
Sullivan, E.G. and A.O. Haugen. 1956. Age determination of foxes by x-ray of forefeet. Journal of Wildlife Management 36: 166-70.
Sutton, R. 1958. A Manitoba record of the Gray Fox. Journal of Mammalogy 39:439-440.
Tate, P., pers. comm. 2014. Telephone conversation with J. Pearce. July 2014. Furbearer Project Leader, New Hampshire Fish and Game, Concord, New Hampshire.
Trapp, G.R. 1978. Comparative behavioural ecology of the ringtail (Bassariscus astutus) and gray fox (Urocyon cineroargenteus) in southwestern Utah. Carnivore 1: 3-32.
Trapp, G.R., and D.L. Hallberg. 1975. Ecology of the grey fox (Urocyon cinereoargenteus): a review. pp. 164-178 in M.W. Fox (ed.). The Wild Canids: their Systematics, Behavioral Ecology and Evolution. Van Nostrand Reinhold Company, New York, NY. xvi + 508 pp.
Tullar, B.F., Jr., and L.T. Berchielli Jr. 1982. Comparison of red foxes and gray foxes in central New York with respect to certain features of behavior, movement and mortality. New York Fish and Game Journal 29: 127-133.
Van den Broeck, J. 2014a. Records of confirmed Grey Fox (Urocyon cineroargenteus) Northwestern Ontario. Unpublished Report by the Ontario Ministry of Natural Resources, Fort Frances, Ontario
Van den Broeck, J. 2014b. Species occurrence record - MNR Fort Frances District, Grey Fox. Unpublished Report by the Ontario Ministry of Natural Resources, Fort Frances, Ontario.
Vermont Department of Fish and Wildlife (VTDFW). 2014. 2014 Vermont Hunting, Fishing and Trapping Laws and Guide: Fish and Wildlife Regulations. Vermont Department of Fish and Wildlife. [Accessed 1 July 20114].
Waters, J.H. 1964. Red fox and gray fox from New England archeological sites. Journal of Mammalogy 45: 307-308.
Waters, J.H. 1967. Foxes on Martha's Vineyard, Massachusetts. Journal of Mammalogy 48: 137-138.
Weston Glenn, J., D. Civitello, and S. Lance. 2009. Multiple paternity and kinship in the gray fox (Urocyon cinereoargenteus). Mammalian Biology 79:394-402.
Weston, J., and L. Brisbin 2003. Demographics of a protected population of gray foxes (Urocyon cinereoargenteus) in South Carolina. Journal of Mammalogy 84:996-1005.
Wilson, D. E., and D. M. Reeder (editors). 2005. Mammal Species of the World: a Taxonomic and Geographic Reference. Third Edition. John Hopkins University Press, Baltimore, MD. xviii + 2142 pp. [Accessed June 2014].
Wintemberg, W.J. 1921. Archaeological evidence concerning the presence of the grey fox (Urocyon sp.) in Ontario. Canadian Field-Naturalist 35: 19-20.
Wintemberg, W.J. 1928. Uren prehistoric village site, Oxford County, Ontario. National Museum of Canada Bulletin No. 51, Anthropological Series No. 10. 97 pp.
Wintemberg, W.J. 1936. Roebuck prehistoric village site, Grenville County, Ontario. National Museum of Canada Bulletin No. 83, Anthropological Series No. 19. 178 pp.
Wintemberg, W.J. 1939. Lawson prehistoric village site, Middlesex County, Ontario. National Museum of Canada Bulletin No. 94, Anthropological Series No. 25. 104 pp.
Wintemberg, W.J. 1948. The Middleport prehistoric village site, Brant County, Ontario. National Museum of Canada Bulletin No. 109, Anthropological Series No. 27. v+ 79 pp.
Wisconsin Department of Natural Resources (WIDNR). 2013. Small game hunting regulations, 2013 fall turkey, and 2014 spring turkey regulations. PDF. [Accessed 1 July 2014].
Wong, J., (and 4 co-authors) Great Lakes Ice Climatology Update: Winter 2006-2011. GLERL-155. [Accessed August 2015].
Wood, J.E. 1958. Age structure and productivity of a gray fox population. Journal of Mammalogy 39: 74-86.
Wood, J.E., D.E. Davis, and E.V. Komarek. 1958. The distribution of fox populations in relation to vegetation in southern Georgia. Ecology 39: 160-162.
Wooding, J.B. 1984. Coyote food habits and the spatial relationship of coyotes and foxes in Mississippi and Alabama. MSc Thesis, Mississippi State University, State College.
Wooding, S., and R. Ward. 1997. Phylogeography and Pleistocene evolution in the North American black bear. Molecular Biology and Evolution 14:1096-1105.
Yearsley, E.F., and D.E. Samuel. 1980. Use of reclaimed surface mines by foxes in West Virginia. Journal of Wildlife Management 44: 729-734.
Yuan Chung Zee. 1999. Rhabdoviridae. pp. 412-417 in D.C. Hirsh, and Yuan Chung Zee (eds.) Veterinary Microbiology. Blackwell Science Inc., Malden, MA. vii + 479pp.
Dr. Jennie L. Pearce was born in Australia and immigrated to Canada in 1999. In both countries her research has focused on modelling the distribution, viability and habitat requirements of wildlife to inform and guide conservation efforts. She has developed spatial distribution models for mammal species with limited distribution records, developed population viability models for species with small populations and modelled the temporal population changes expected to occur when landscapes are modified due to human activities. She has written or co-authored 10 COSEWIC reports and has published more than 40 scientific papers in the area of conservation biology.
No collections were examined.
| Date | Location | Observer | Source |
|---|---|---|---|
| 1946/1947 | Sprague | A. Suchrov | Sutton 1958 |
| 12 Jan 1957 | St. Adolphe | M. Bouchard | MMMN records; Sutton 1958 |
| 9 Dec 1977 | 7mi S, 2mi W of Hadashville | E. Shindler | MMMN records |
| 20 Oct 2000 | 5mi N of Piney | P. Krahn | MMMN records |
| Approx 2002 | Whiteshell (trapped) | - | Berezanski pers comm. |
| Every couple years | Sprague area (roadkill) | - | Berezanski pers comm. |
| Individuals | Date | Location | Source (Observer) | NHIC observation number |
|---|---|---|---|---|
| 1-2 | 1 Jan 1944 2 Oct 1944 | Rainy River Co., Wild Potato Lake (ROM16311) | ROM records (S. Boy) | 219386, 219387 |
| 1-2 | 22 Feb 1982 | Thunder Bay Co., just N and W of Shebandowan Lake | M. Lankester, pers. comm. (V. Hollet) | 219385, 219565 |
| 1-2 | 10 Dec 91 | Thunder Bay Co., S of Dog Lake | B. Wall, pers. comm. (B. Wall) | 219384, 219383 |
| 1 | Dec 1997 | Rainy River Co., 5km SE of Atikokan | Trapped, B. Jackson, pers. comm. (a trapper) | 219564 |
| 1 male | Feb 2006 | Northwest of Fort Frances, 800m south of Little Pine Lake | Trapped, Van den Broeck (2014a) | - |
| 1 male | 2008a | Rocky Inlet | Trapped, Van den Broeck (2014a) | - |
| 1 male | 2008b | Couchiching | Trapped, Van den Broeck (2014a) | - |
| 1 | Nov 2010 | Mountain Lake Rd., Thunder Bay | Photo, Van den Broeck (2014a) | - |
| 3 | 11 Feb 2011 | Fern Rd., Whitefish Lake Thunder Bay | Photo, Van den Broeck (2014a) | - |
| 1 | 9 Feb 2011 | Calm Lake, Atikokan | Photo, Van den Broeck (2014a) | - |
| 1 | 5-7 May 2012 | Sibley Peninsula, Thundercape Bird Observatory | Observed, Van den Broeck (2014a) | - |
| 1 | 2012 | Mine centre, Fort Francis District | Trapped, Van den Broeck (2014a) | - |
| 2 | Winter 2012 | Mountain Rd, Thunder Bay | Photo, Van den Broeck (2014a) | - |
| 4 | Summer/fall 2013 | Mountain Rd., Thunder Bay | Observed, then one road-killed, Van den Broeck (2014a) | - |
| 1 Male | Nov. 2013 | Delints Rd - near White Fish Lake area | Trapped, Van den Broeck (2014a) | - |
| 1 male | Winter 2014 | Dorion | Observed then died, Van den Broeck (2014a) | - |
| 1 male, 1 female | 2014 | Rainy River, Fort Francis District | Roadkill, Van den Broeck (2014a) | - |
| 1 male | 2014 | Thunder Bay, Devon Rd - near border | Trapped, Van den Broeck (2014a) | - |
| 1 lactating female | Summer 2015 | Mountain Rd, Thunder Bay | B. Ratcliff (photo) | - |
| Individuals | Date | Location | Source (Observer) | NHIC observation Number |
|---|---|---|---|---|
| 1 - 4 | Oct 1983 | Essex Co., Pelee Island at the highest elevation | Harris 1994 (L. Defraeve) | 79378 |
| 1 | Oct 1983 | Essex Co., Pelee Island at the N point | Harris 1994 (L. Defraeve) | 79379 |
| 1 | 1 Nov 1984 | Essex Co., Pelee Island, Stone Rd. Alvar (ROM91445) | ROM records (T. Gardner) | 79386 |
| 1 | Apr 1985 | Essex Co., Pelee Island, dead on road | D. Coulson, pers. comm. (D. Coulson, C. Campbell) | 79377 |
| 1 | 31 Jan 1989 | Essex Co., Pelee Island, south half | Harris 1994 (K. Hillman) | 79385 |
| 1 | around 1990 | Essex Co., Pelee Island | N. Beattie, pers. comm. (N. Beattie) | 79376 |
| 1 | 1994 | Essex Co., Pelee Island, at N quarry site | B. Porchuk, pers. comm. (B. Porchuk) | 79375 |
| 2 | Nov 1996 | Essex Co., Pelee Island, on road at Fish Point Nature Reserve (2 individuals) | B. Porchuk, pers. comm. (B. Porchuk) | 19374 |
| 1 | Spring 1997 | Essex Co., Pelee Island, on B. Porchuk's property on Curry Dyke Rd. at S end of island | B. Porchuk, pers. comm. (B. Porchuk) | 79384 |
| 1 | 1 Jun 1998 | Essex Co., Pelee Island | London Free Press 1998 (D. Hodare) | 79383 |
| 1 | 7 Jun 1998 | Essex Co., Pelee Island, on road at Fish Point Nature Reserve | P. Pratt, pers. comm. (P. Pratt) | 79382 |
| 1 | 12 Apr 2000 | Essex Co., Pelee Island, Fish Point Nature Reserve | B. Porchuk, pers. comm. (B. Porchuk) | 79381 |
| 1 | 11 Jun 2000 | Essex Co., Pelee Island, crossing W Shore Rd. north of ferry dock | P. Pratt, pers. comm. (Essex Co. Field Naturalists Club) | 79380 |
| 1 | Jan 2001 | Essex Co., Pelee Island | N. Beattie, pers. comm. (N. Beattie) | 79373 |
| 1 | 17 June 2010 | NCC Cabin, Pelee Island, Middle Point Woods (NCC) | Sam Brinker (Photograph) | 219350 |
| 1-16 | Winter 2012/2013 | 8 locations on Pelee Island | Bowman et al. (2013) | - |
| 1 | Dec. 2012 | Road killed, Pelee island | C. Sadowski, pers. comm. | - |
| Individuals | Date | Location | Source (Observer) | NHIC observation Number |
|---|---|---|---|---|
| 1 | Pre-1900 | Hillman Marsh | - | 219549 |
| 1 female | 20 Feb 1948 | Port Colborne | - | 219391 |
| 1 male | 26 Mar 1949 | Niagara Co., St. Catharines, Grantham Geo. Twp. (ROM19592) [NW Niagara falls] | ROM records (B. Demars) | 219396 |
| 1 female | 1 Jan 1952 | Niagara Co., Wainfleet Twp. (near Chamber's Corners) (ROM21489) | ROM records (A.H. Muma) | 219390 |
| 1 male | 2 Jan 1952 | Niagara Co., Fort Erie Twp. (ROM21491) | ROM records (Ont. Dept. Lands & Forests) | 219393 |
| 1 female | 15 Nov 1952 | Niagara Co., Port Colborne, Humberstone Geo. Twp. (ROM22501) | ROM records (A.H. Muma) | 219392 |
| 1 female | 1 Jan 1954 | Niagara Co., Niagara Falls Twp., near Chippawa (ROM25502) | ROM records (Ont. Dept. Lands & Forests) | 219395 |
| 1 | Jul 1964 | Niagara Co., Grimsby Twp., near Spring Creek rd. and Park rd. near dump | G. Meyers, pers. comm. | 219397 |
| 1 | 1965 | Haldimand-Norfolk Co., Grand River Marshes, SE of Dunnville | Gartshore 1987 (W. Houser) | 219388 |
| 1 | Jan 1978 | Simcoe Co., near Stayner | Dobbyn 1994 (local trapper) | 219351 |
| 1 male | 11 Oct 1978 | Lot 15, Concession 2 Goderich Township, Huron County | Specimen # 8344, OMNR Rabies Surveillance data | - |
| 1 | 25 Oct 1978 | Haldimand-Norfolk Co., Norfolk Twp., 6km N of Cultus, Conc. VIII Walsingham (ROM82842) | ROM records (O. Wilson) | 219545 |
| 1-2 | 1978/79 trapping season | Niagara Co., N of Allanburg | Harris 1994 (J. Caldwell) | 219569 |
| 1 | 21 Mar 1979 | 2km SW of Wiarton | - | 219563 |
| 1 female | 10 Nov 1979 | Lot 16, Concession 3, Nottawasaga Township, Lambton County | OMNR Rabies Surveillance data | - |
| 1 | 1979/80 trapping season | Niagara Co., Thorold Twp., trapped in what is now the Brock Industrial Park | Harris 1994 (L. Schmaltz) | 219568 |
| 1 | 1980 (and seen from 1975-1980) | Haldimand-Norfolk Co., Haldimand Twp., Oriskany Sandstone and Woodlands, 7km W of Cayuga, ca. 1.5km S of Clanbrassil | MacDonald 1989 (S. Banticks) | 219389 |
| 1 male | 15 Oct 1980 | Lambton Co., Enniskillen Twp., about 4km E of Petrolia, Lot 26, Concession 12 | Bachmann and Lintack 1982 (W. Lintack) | 219550 |
| 1 male | 1 Dec 1980 | Lot 34, Concession 8, Lindsay Township, Bruce County | Specimen # 13577 in OMNR Rabies Surveillance data | - |
| 1 | 15 Oct 1980 | Essex Co., approx. 0.5-1mi N or Cottam and 4mi E of Essex | Harris 1994 (P. Jacques) | 219552 |
| 1 | 1980 1985 | Essex Co., Dominion Blvd. in Windsor | P. Pratt, pers. comm. (Gary's taxidermy) | - |
| 1 male | 1 Apr 1982 | Lot 29, Concession 13, Enniskillen, Lambton County | Specimen # 15788, OMNR Rabies Surveillance data | - |
| 1 | 1982 | Haldimand-Norfolk Co., Delhi Twp., N of Waterford Ponds on Concession Rd. VI | Gartshore 1987 (R. Bergman) | 219548 |
| 2 | 23 May 1984 | Essex, Co., Windsor, north side of Todd Lane, LaSalle/Windsor boundary (2 adults) | P. Pratt, pers. comm. (S. Wardle, P. Pratt) | 219381 |
| 1-2 | 4 June 1984 | Edge of woodlot behind suburban house, Near Ojibway Prairie, Windsor | P. Pratt (photo) | 219353 |
| 1 | 1985 | Essex Co., Windsor, southern edge of Spring Garden ANSI | P. Pratt, pers. comm. (S. Wardle) | 219382 |
| 1 | 27 Oct 1985 | Essex Co., Anderton Twp., Amherstburg, Lot 42, Concession 1 | Harris 1994 (E. Meloche) | 219553 |
| 1 | 10 June 1986 | Medonte (Coldwater), Simcoe County | Specimen ID #42138, CFIA submission # 86RABN05294 | - |
| 1 | 29 Jul 1986 | Waterford, Norfolk County | Specimen ID #22074, CFIA submission # 86RABN05294 | - |
| 1 | Nov 1986? | Essex Co., Anderton Twp., Amherstburg, Lot 40, Concession 1 | Harris 1994 (E. Meloche) | 219556 |
| 1 | Nov 1987? | Essex Co., Anderton Twp., Amherstburg, Lot 40, Concession 1 | Harris 1994 (E. Meloche) | 219557 |
| 1 | Nov/Dec 1987 | Essex Co., Anderton Twp., Amherstburg, Lot 42, Concession 1 | Harris 1994 (E. Meloche) | 219558 |
| 1 | Nov/Dec 1987 | Essex Co., Anderton Twp., Amherstburg, Lot 42, Concession 1 | Harris 1994 (E. Meloche) | 219559 |
| 1-2 | Dec 1989 or Jan 1990 | Lambton Co., Sombra Twp., between Concessions 7 and 8 | Harris 1994 (G. English) | - |
| 1 | before 19906 | Essex Co., Mersea Twp., Hillman Marsh, observed mounted specimen | P. Pratt, pers. comm. (P. Pratt) | - |
| 1 | 14 Nov 19906 | Simcoe Co., Mara Twp., Lot 26, Concession 11 | Harris 1994 (B. Edinger) | - |
| 1 | 9 May 1991 | Niagara Co., 7 Bedford Park, Grimsby | G. Meyers, pers. comm. (G. Meyers) | 219398 |
| 1 | 7 Feb 1997 | Wellington Co., Nichol Twp., ca. 3km N of Elora in woodlot near Irvine Creek | D. Coulson, pers. comm. (D. Coulson) | 219561 |
| 1 female | 28 Oct 2001 | Lot 13, Concession 2, Goderich, Huron County | Specimen # 74309, OMNR Rabies Surveillance data | - |
| 1 | 19 Aug 2002 | South Cameron Road, west of Dougall St, Windsor | - | pending |
| 1 | Unknown | 1.5km SE of Craighurst, carcass collected | - | 219405 |
| Individuals | Date | Location | Source (Observer) | NHIC observation Number |
|---|---|---|---|---|
| 1 | Jan 1942 | Stormont, Dundas & Glengarry Co., Kenyon Twp., 4mi W of Alexandria (ROM15707) | - | 219406 |
| 1 | Feb 1944 | Frontenac Co., Kenebec Twp., 6mi E of Kaladar (ROM16214) | ROM records (R. Knight) | 219401 |
| 1 female | 20 Feb 1948 | Niagara Co., Port Colborne, Humberstone Geo. Twp. (ROM19494) | ROM records (W. Lee) | 219391 |
| 1 female | 27 Apr 1948 | Leeds & Grenville Co., Escott Twp. (ROM18801) | ROM records (B. Reed) | 219570 |
| 1 | 1 Jul 1951 | Leeds & Grenville Co., Elizabethtown Twp., near Brockville (ROM21579) | ROM record (Unknown) | 219399 |
| 1 male | 27 Nov 1952 | Lennox & Addington Co., Abinger Twp. (ROM22493) | ROM records (I. Brown) | 219571 |
| 1 | 1963 | Wellington Co., on the road between Breslau and Guelph | Campbell et al. 1972 (Unknown) | 219562 |
| 1 female | 19 Mar 1977 | Peterborough Co., 3km NE of Cavan (ROM80081) | ROM record (RD McRae) | 219380 |
| 1 | Jan 1978 | Lennox & Addington Co., on Wolfe Island near Point Alexander, Concession 6 | Harris 1994 (R. Fawcett) | 219567 |
| 1 | 31 Jan 1979 | Lennox & Addington Co., Holiday Point, Wolfe Island | Harris 1994 (R. Hodge) | 219566 |
| 1 male | 24 Oct 1979 | Durham Co., Pickering Twp., near Balsam, 5km E of Claremont (ROM82841) | ROM records (P. Hall) | 219394 |
| 1 | 31 May 1979 | North of Tar Island, approx 2mi down river from the Thousand Island Bridge | - | 219400 |
| 1 | Nov 1980 | Renfrew Co., Lyell Twp. | Harris 1994 (B. Coulas) | - |
| 1 | Jan 1981 | Lanark Co., Pakenham Twp., Concession 12 | Harris 1994 (C. Coe) | - |
| 1 | Between 1983 and 1985 | Northumberland Co., Cramahe Twp. | Harris 1994 (B. Stratton) | - |
| 1 male | 15 Dec 2000 | Lanark Co., Drummond Twp., along Hwy 7 near Perth | Specimen # 68215, OMNR Rabies Surveillance data | 219402 |
| 1 | Oct 2008 | St. Lawrence Islands Parkway | Roadkill, MNRF (L. Bruce) | - |
| 1 | 5 March 2014 | Crossing Hwy 7, Concession 9, Lanark County | Observed, MNRF (S. Thompson) | - |
| 1 | Nov/Dec 2014 | Mills St in Bishop Mills | Roadkill, MNRF (R. Ciraco) | - |
| Individuals | Date | Location | Source (Validated by) |
|---|---|---|---|
| 1 | 1975 | Saint-Bernard-de-Lacolle | Trapped, M. Richard (H. Jolicoeur); S. Thérien, K. Racey |
| 1 male | Feb 1981 | Saint-Bernard-de-Lacolle | Trapped, P. Martin (H. Jolicoeur) |
| 1 female | Winter 1984-1985 | Saint-Bernard-de-Lacolle | Trapped, P. Martin (H. Jolicoeur) |
| 1 male | 1990-2000 | Saint-Anicet | Trapped, A. Lebrun/P. Martin (H. Jolicoeur) |
| 1 female | 1990-2000 | Saint-Anicet | Trapped, A. Lebrun/P. Martin (H. Jolicoeur) |
| 1 | 23 Oct 2003 | Saint-Stanislas-de-Kostka | Trapped, A. Lebrun/P. Martin (H. Jolicoeur) |
| 1 male | 15 Dec 2007 | St-Rémi-de-Napierreville | Trapped, D.Tremblay/P. Martin (H. Jolicoeur) |
| 1 | 2009 | Hemmingford | Observed, S. Thérien/P. Martin (H. Jolicoeur) |
| 1 | 2009 | Hemmingford | Observed, S. Thérien/P. Martin (H. Jolicoeur) |
| 1 | 8 Nov 2010 | Saint-Bernard-de-Lacolle | Livetrapped, P. Martin (H. Jolicoeur) |
| 1 female | 2012 | Saint-Bernard-de-Lacolle | Roadkill, P. Martin (G. Tremblay) |
| 1 male | 30 Nov 2012 | Saint-Bernard-de-Lacolle | Trapped P. Martin (G. Tremblay) |
| 1 male | 2013 | Hemmingford | Trapped P. Martin (G. Tremblay) |
| 1 | 12 Jan 2012 | Léry | Trapped S. Lessard [APF] (E. Dalpé-Charron/F. Landry) |
| 1 | 2012 | Saint-Bernard-de-Lacolle | Trapped, C. Lemay [APF] (E. Dalpé-Charron/F. Landry) |
| 1 | 28 Nov 2013 | Hemmingford | Trapped, P. Martin (E. Dalpé-Charron/F. Landry) |
| 1 | 5 Jan 2014 | Hemmingford | Trapped, S. Richard (E. Dalpé-Charron/F. Landry) |
| 1 | 11 Feb 2014 | Hinchinbrooke | Trapped, A. Lebrun (E. Dalpé-Charron/F. Landry) |
| 1 | 2014 | La Prairie | Video (P. Blais) |
| 1 | 2014 | La Prairie | Video (P. Blais) |
| Individuals | Date | Location | Source (Validated by) |
|---|---|---|---|
| 1 | 1894 | Cookshire-Eaton (Johnville) | Observed, K Racey (H. Jolicoeur) |
| 1 juvenile female | 12 Dec 2011 | Sherbrooke | Roadkill, M-C Benoît (G. Tremblay) |
| 1 | 8 Feb 2012 | Cookshire-Eaton | Trapped, M. Rolfe (E. Dalpé-Charron/F. Landry) |
| 1 | 1 Jan 2014 | Saint-Malo | Trapped, M. Blais (E. Dalpé-Charron/F. Landry) |
| 1 | 5 Dec 2013 | Sherbrooke | Trapped, M. Grondin (E. Dalpé-Charron/F. Landry) |
| Individuals | Date | Location | Source (Validated by) |
|---|---|---|---|
| 1 | 1940-1950 | Saint-Augustin-de-Woburn | Trapped (H. Jolicoeur) |
| 1 | 19 Dec 2011 | Lac-Mégantic | Roadkill?, A. Gauthier [APF] (E. Dalpé-Charron/F. Landry) |
| 1 | 12 Feb 2013 | Frontenac | Trapped, Y. Poirier (E. Dalpé-Charron/F. Landry) |
| 1 | 14 Nov 2013 | Saint-Augustin-de-Woburn | Trapped, A. Bédard (E. Dalpé-Charron/F. Landry) |
| Individuals | Date | Location | Source (Validated by) |
|---|---|---|---|
| 1 | 1 Dec 1986 | Saint-Benjamin | Trapped, G & F Caron (H. Jolicoeur) |
| 1 adult male | Autumn 1996 | Sainte-Justine | Trapped, E. Houle (H. Jolicoeur) |
| 1 adult male | 15 Nov 2005 | Sainte-Aurélie | Trapped J-F Caouette/G. Fortin (H. Jolicoeur) |
| 1 | Dec 2006 | Saint-Côme-Linière | Trapped, G. Poulin (H. Jolicoeur) |
| 1 adult male | 2007 | Saint-Benoît-Labre | Trapped, B. Pouliot (H. Jolicoeur) |
| 1 adult male | 22 Dec 2014 | Voir Griffe | Trapped, E. Dalpé-Charron |
| 1 male | 20 Nov 2014 | Montmagny | Trapped, E. Dalpé-Charron |
| Individuals | Date | Location | Source |
|---|---|---|---|
| 1 sub-adult male | 29 October 2007 | Rollingdam, Charlotte County | Trapped, (McAlpine et al. 2008) |
| 1 | 19 September 2014 | Oromocto Lake, York County | Trail camera, (McAlpine et al. submitted) |
| Threat Impact | Threat Impact (descriptions) | Level 1 Threat Impact Counts: high range |
Level 1 Threat Impact Counts: low range |
|---|---|---|---|
| A | Very High | 0 | 0 |
| B | High | 1 | 1 |
| C | Medium | 0 | 0 |
| D | Low | 0 | 0 |
| - | Calculated Overall Threat Impact: | High | High |
| Threat Number | Threat | Impact (calculated) |
Impact (calculated) Description |
Scope (next 10 Yrs) |
Severity (10 Yrs or 3 Gen.) |
Timing | Comments |
|---|---|---|---|---|---|---|---|
| 1 | Residential & commercial development | - | - | - | - | - | No evidence of threat |
| 2 | Agriculture & aquaculture | - | - | - | - | - | No evidence of threat. Potential Limiting Factor (same with agriculture and possibly fire) |
| 3 | Energy production & mining | - | - | - | - | - | No evidence of threat |
| 4 | Transportation & service corridors | - | Not a Threat | Pervasive (71-100%) |
Neutral or Potential Benefit | High (Continuing) |
- |
| 4.1 | Roads & railroads | - | Not a Threat | Pervasive (71-100%) |
Neutral or Potential Benefit | High (Continuing) |
* Not likely, but unknown if road kill is driving decline in population * Over the EO, road kill likely limits expansion; if only addressing known breeding population, Pelee Island constitutes where most of the population is impacted (>30%) Members on the threats call believe that the severity actually 'degrades/reduces the affected occurrences' as opposed to having a 'neutral or potential benefit' |
| 5 | Biological resource use | B | High | Pervasive - Large (31-100%) |
Serious (31-70%) |
High (Continuing) |
- |
| 5.1 | Hunting & collecting terrestrial animals | B | High | Pervasive - Large (31-100%) |
Serious (31-70%) |
High (Continuing) |
what % of those animals are exposed: they are all exposed * there has been bycatch taken as far north as North Bay * Pelee Island: has more road kills than trapping * QC: fairly heavily trapped animal * given uncertainty, range chosen 'Pervasive-Large' Species can easily expand if they aren't trapped * species has high mortality rate in US (due to harvest, roadkill) but population maintained by high fecundity |
| 5.3 | Logging & wood harvesting | - | - | - | - | - | Potential Limiting Factor (same with agriculture and possible fire) * species does not like it too open |
| 6 | Human intrusions & disturbance | - | - | - | - | - | No evidence of threat |
| 7 | Natural system modifications | - | - | - | - | - | - |
| 7.1 | Fire & fire suppression | - | - | - | - | - | Potential Limiting Factor (same with agriculture and possibly fire) |
| 8 | Invasive & other problematic species & genes | - | Unknown | Pervasive (71-100%) |
Unknown | Moderate (Possibly in the short term, < 10 yrs) |
- |
| 8.1 | Invasive non-native/alien species | - | Unknown | Pervasive (71-100%) |
Unknown | Moderate (Possibly in the short term, < 10 yrs) |
Canine distemper considered as non-native * what % could be exposed to this disease: all of it * did we ever have a gray fox turned in that died of canine distemper: no |
| 8.2 | Problematic native species | - | Unknown | Large (31-70%) |
Unknown | High (Continuing) |
Rabies considered native * Timing given as "High" based on 2 reported mortality cases in ON |
| 9 | Pollution | - | - | - | - | - | No evidence of threat |
| 10 | Geological events | - | - | - | - | - | No evidence of threat |
| 11 | Climate change & severe weather | - | - | - | - | - | Warming could be a benefit for this species, based on recent range expansion northward in the US |
Classification of Threats adopted from IUCN-CMP, Salafsky et al. (2008).
Page details
- Date modified: