
Béluga (Delphinapterus Leucas) évaluation et rapport de situation du COSEPAC : chapitre 8
Identification des populations
- Population de l’estuaire du Saint-Laurent
- Population de la baie d’Ungava
- Population de l’est de la baie d’Hudson
- Population de l’ouest de la baie d’Hudson
- Population de l’est du haut Arctique et de la baie de Baffin
- Population de la baie Cumberland
- Population de l’est de la mer de Beaufort
- Incertitudes concernant l’identification des populations de bélugas au Canada
Par le passé, aux fins de gestion, les chercheurs ont distingué sept populations de bélugas qui sont présentes dans les eaux canadiennes au moins une partie de l’année (figure 2), soit : 1) la population de l’estuaire du Saint-Laurent (Pippard, 1985); 2) la population de la baie d’Ungava; 3) la population de l’est de la baie d’Hudson (Reeves et Mitchell, 1989; MPO, 2001); 4) la population de l’ouest de la baie d’Hudson (Richard, 1993); 5) la population de la baie Cumberland (Richard et Orr, 1991; MPO, 2002a); 6) la population de l’est du haut Arctique et de la baie de Baffin (Doidge et Finley, 1993); 7) la population de l’est de la mer de Beaufort (MPO, 2000).
Figure 2 : Localisation des sept populations canadiennes de bélugas : 1) population de l’estuaire du Saint-Laurent; 2) population de la baie d’Ungava; 3) population de l’est de la baie d’Hudson; 4) population de l’ouest de la baie d’Hudson; 5) population de l’est du haut Arctique et de la baie de Baffin; 6) population de la baie Cumberland; 7) population de l’est de la mer de Beaufort
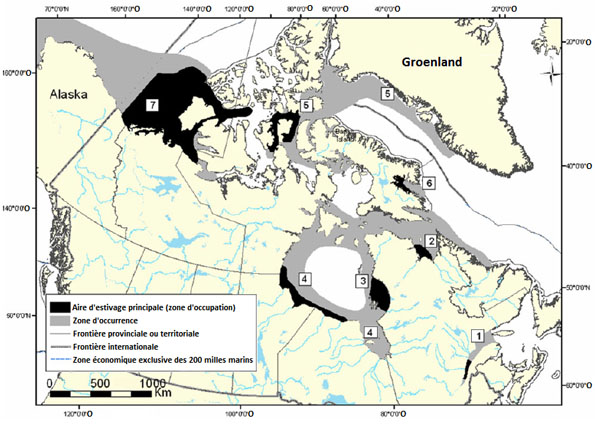
Données modifiées par rapport à celles du MPO, 2002b.
Selon cette division, les populations canadiennes de bélugas se rassemblent toutes dans des estuaires pendant la saison des eaux libres. Dans la plupart des cas, chacune a son propre territoire estival sur les côtes et au large. En automne, en hiver et au printemps, les aires de répartition sont souvent contiguës ou se chevauchent.
Les premières données ayant mené à la reconnaissance de différentes populations de bélugas portaient sur les différences de taille entre bélugas de diverses régions de l’Arctique (Sergeant et Brodie, 1969). À l’issue d’un réexamen des données et des méthodes d’analyse, il a été possible de confirmer l’existence de différences mineures, mais celles-ci n’ont pas été jugées suffisantes pour être utilisées de façon pratique comme critère pour différencier des populations de bélugas voisines les unes des autres (Doidge, 1990). Stewart (1994) a découvert d’importantes différences de taille entre plusieurs populations bien séparées sur le plan géographique, mais il a également signalé que les différences de taille constituent aujourd’hui un paramètre moins fiable pour l’identification des populations, surtout avec les nouvelles techniques de discrimination génétique des populations.
Les chercheurs ont été plus nombreux à accepter la discrimination fondée sur les aires d’estivage, qu’ils pensaient être des zones principalement côtières renfermant des lieux comme les estuaires où se rassemblent un grand nombre d’individus (Sergeant et Brodie, 1975; Smith et al., 1985). Cette hypothèse a été corroborée par des études sur le comportement du béluga dans des estuaires, qui ont révélé la philopatrie de ce cétacé et sa grande fidélité à certains sites précis (Caron et Smith, 1990; Smith et al., 1994). À l’un de ces estuaires, on a observé que des individus reconnaissables à des cicatrices distinctives retournaient fréquemment et rapidement dans l’estuaire peu après les chasses (Caron et Smith, 1990).
Depuis peu, les analyses génétiques, aussi bien de l’ADN mitochondrial (ADNmt) que de l’ADN microsatellite, sont utilisées de plus en plus couramment pour différencier les populations de bélugas (de March et al., 2001 et 2002). Les scientifiques se rendent progressivement compte de l’existence d’un certain nombre de problèmes complexes associés à la représentativité des animaux échantillonnés, à la portée et à l’étendue des analyses génétiques elles-mêmes et à leur normalisation d’une étude à l’autre, ainsi qu’aux méthodes statistiques qui conviennent pour l’analyse d’ensembles de données génétiques complexes (de March et al., 2002; Palsbøll et al., 2002; de March et Postma, 2003). Ces études génétiques confirment l’existence de populations déjà reconnues, mais elles révèlent aussi l’existence de nouvelles populations. D’autres types d’études ont été réalisées dans le but de circonscrire la période de migration et les parcours employés. Celles-ci faisaient appel au pistage satellitaire (Martin et al., 1993), aux connaissances traditionnelles détaillées des Autochtones et à d’autres indicateurs tels que la signature de contaminants (Innes et al., 2002a).
Le présent rapport vise à définir des unités désignables en fonction de l’aire de répartition géographique et de différences génétiques (COSEPAC, 2003 - Annexe F5). Ces dernières années, de nouvelles études génétiques ont révélé l’existence d’un plus grand nombre de populations de bélugas qu’on avait imaginé. Cependant, en raison de lacunes au chapitre de l’échantillonnage, il n’a pas encore été possible de les définir avec précision ou de façon pratique aux fins de gestion. Comme aucune de ces nouvelles données génétiques ne contredit l’existence des populations reconnues, qui ont toutes reçu un statut, ce sont ces populations qui formeront l’assise et le point de départ du présent examen. Au besoin, le présent rapport fait mention de toute preuve solide concernant le profil génétique, l’aire de répartition géographique, la migration ou d’autres caractéristiques écologiques qui plaide en faveur de la désignation d’autres unités évolutionnaires significatives (Waples, 1991).
Les bélugas de l’estuaire du Saint-Laurent sont considérés depuis longtemps comme une population relique de l’Arctique. Le caractère distinct de cette petite population de bélugas, concentrée principalement aux environs de l’embouchure du Saguenay, tient à l’absence d’un nombre important de bélugas dans les secteurs avoisinants. Seuls quelques individus fréquentent la côte nord du fleuve Saint-Laurent ou la côte sud du Labrador, ce qui confirme l’absence d’un lien encore existant entre les bélugas du Saint-Laurent et les populations du littoral sub-arctique du Québec (Reeves et Mitchell, 1984; Michaud et al., 1990; Curren et Lien, 1998).
Les bélugas du fleuve Saint-Laurent sont plus distincts sur le plan génétique que toutes les autres populations canadiennes de bélugas. Ils possèdent un haplotype qui ne se retrouve nulle part ailleurs et un autre qu’ils partagent uniquement avec les bélugas de l’est de la baie d’Hudson, en particulier ceux de l’estuaire de la rivière Nastapoka. La population géographiquement isolée du Saint-Laurent et la population de l’est de la baie d’Hudson sont celles qui présentent la plus faible diversité d’haplotypes d’ADNmt et d’allèles microsatellites (de March et al., 2002). Il se peut que les bélugas du Saint-Laurent et de l’est de la baie d’Hudson soient issus de la mer intérieure créée par les lacs Agassiz et Ojibway, refuge de la période de glaciation wisconsinienne. Plus récemment, de March et al. (2002) ont émis l’hypothèse voulant que les bélugas de l’est de la baie d’Hudson aient été les premiers à quitter leur refuge de l’Atlantique après la disparition de la première barrière de glace il y a quelque 8 000 ans avant le présent (Fulton, 1989). Les autres populations canadiennes, y compris les autres populations de l’est de l’Arctique et celle de l’ouest de la baie d’Hudson, seraient issues d’un refuge situé dans l’ouest (Brown, 1996; Brown-Gladden et al., 1997; Brown-Gladden et al., 1999). Il faudra réaliser d’autres études, en particulier sur les bélugas de l’ouest de la Russie, pour élucider le mystère de la dispersion postglaciaire de l’espèce.
La population dite de la baie d’Ungava était considérée comme distincte en raison de son aire de répartition estivale, qui était concentrée principalement près de l’estuaire de la rivière Mucalic, même si l’on sait qu’un faible nombre d’individus se concentraient dans de petits estuaires comme ceux de la rivière George et de la rivière de la Baleine. Cette population, qui n’a jamais compté un effectif nombreux (Reeves et Mitchell, 1989), a été considérablement réduite par l’exploitation commerciale et par la chasse de subsistance. La rivière Mucalic n’est plus une aire de rassemblement importante pour les bélugas (données inédites de T. Smith, obs. pers., 1985). En raison du faible nombre d’animaux capturés dans ce secteur ces dernières années, nous ne possédons que quelques échantillons pour l’étude de la structure génétique de cette population. À partir d’un petit échantillon de bélugas (n=11) prélevé dans la région de la baie d’Ungava, Mancuso (1995) a constaté une diversité d’haplotypes plus élevée que prévu, compte tenu du faible effectif de la population (Smith et Hammill, 1986). Cette découverte a été corroborée par d’autres échantillonnages et d’autres analyses (de March et Maiers, 2001) qui ont révélé une forte diversité d’haplotypes chez 33 bélugas capturés à plusieurs endroits dans la baie d’Ungava; cependant, certains de ces animaux ont été abattus hors de l’aire de répartition principale de cette population et pourraient donc appartenir à d’autres populations. Cette forte diversité génétique pousse les chercheurs à se demander si les bélugas de la baie d’Ungava font partie d’une ou de plusieurs autres populations. Comme les principaux lieux d’estivage, tels que l’estuaire de la rivière Mucalic, sont maintenant presque inoccupés, il se peut que nous ne sachions jamais si les fréquences alléiques chez les bélugas de la baie d’Ungava étaient différentes de celles des populations d’autres régions de l’Arctique.
Toutes les données sur les caractéristiques génétiques, la répartition géographique et l’abondance révèlent que cette population a atteint des niveaux indétectables ou qu’elle est disparue.
Jusque dans les années 1950, les bélugas étaient relativement nombreux en été le long de la côte nord du Labrador, mais ils s’y font très rares depuis (Brice-Bennett, 1978). On signale chaque année à l’association des Inuits du Labrador environ une douzaine d’observations estivales et deux ou trois captures faites en été (J. Rowell, Labrador Inuit Association, comm. pers., 2004). En raison de leur proximité géographique, ces bélugas pourraient avoir eu des affinités avec la population de la baie d’Ungava.
À l’origine, cette population était définie par sa forte concentration dans les estuaires de la rivière Nastapoka et de la Petite rivière de la Baleine, son principal territoire d’estivage côtier s’étendant de Kuujjuarapik à Inukjuak. Des recensements aériens entrepris en 1983 ont révélé que les bélugas étaient aussi répartis dans la région vers le large jusqu’aux îles Belcher (Smith et Hammill, 1986; Kingsley, 2000; Gosselin et al., 2002).
Les analyses d’ADN mitochondrial montrent qu’il existe une différence génétique entre les bélugas de l’est et de l’ouest de la baie d’Hudson (Mancuso, 1995; Brennin et al., 1997; Brown-Gladden et al., 1997; de March et Postma, 2003). D’autres analyses d’ADN mitochondrial et de microsatellites ont révélé que les bélugas des îles Belcher sont systématiquement différents des populations de l’est et de l’ouest de la baie d’Hudson. Ces bélugas pourraient donc former une population distincte ou un mélange constant d’au moins deux populations (de March et Postma, 2003).
Dans le cas de la population de l’est de la baie d’Hudson, la limite sud de l’aire de répartition n’est pas clairement définie. À Sanikiluaq, dans les îles Belcher, près de l’aire d’estivage des bélugas de l’est de la baie d’Hudson, les chasseurs capturent des individus qui semblent issus d’une population distincte ou d’un mélange constant d’autres populations. De nombreux bélugas passent l’été dans la baie James, secteur qui touche à l’extrémité sud du territoire de la population de l’est de la baie d’Hudson et de Sanikiluaq. Des bélugas occupent également le littoral du nord de l’Ontario, de la baie James jusqu’à Churchill, au Manitoba. Les rares échantillons génétiques de ces animaux voisins qui ont été analysés indiquent un lien de parenté avec la population de l’ouest de la baie d’Hudson (B. de March, comm. pers.). De récents travaux de marquage des bélugas dans le fleuve Nelson, au Manitoba, révèlent que les bélugas de la population de l’ouest de la baie d’Hudson fréquentent eux aussi le littoral du nord de l’Ontario et qu’ils se rendent jusque dans la baie James (P. Richard, comm. pers.).
Les bélugas capturés pendant la chasse printanière et automnale le long de la côte sud du détroit d’Hudson présentent différents génotypes, tout comme le faible nombre de bélugas récoltés dans la baie d’Ungava. Ces régions accueillent des bélugas de l’est de la baie d’Hudson (Bourdages et al., 2002), de l’ouest de la baie d’Hudson et du sud-est de l’île de Baffin en raison du régime favorable des glaces en hiver juste au nord de la baie d’Ungava et dans les zones extracôtières du détroit d’Hudson (de March et Postma, 2003).
Il semble exister de solides données génétiques pour étayer le statut de population distincte qui a été conféré aux bélugas de l’est de la baie d’Hudson. Les chercheurs disposent également d’un volume croissant de données qui corroborent l’hypothèse selon laquelle des individus de cette population sont capturés dans d’autres collectivités du Nunavik, parfois dans des régions aussi éloignées que la baie d’Ungava.
Les bélugas de cette population présentent un grand nombre des haplotypes que partagent également les autres populations canadiennes. Cependant, les haplotypes les plus courants chez les populations de l’est de la baie d’Hudson et de l’estuaire du Saint-Laurent, de même que certains haplotypes présents chez les bélugas de la mer de Beaufort, sont relativement rares chez les individus de l’ouest de la baie d’Hudson (de March et Postma, 2003). Des échantillons génétiques de bélugas de l’ouest de la baie d’Hudson ont été prélevés dans des collectivités aussi éloignées les unes des autres que Churchill (Manitoba), Sanikiluaq, Repulse Bay et les Territoires du Nord-Ouest, et ils présentent une grande diversité d’haplotypes et de microsatellites. Ce groupe, dans son ensemble, est donc moins homogène sur le plan génétique que celui de l’est de la baie d’Hudson. Par exemple, dans des études récentes, seuls 75 spécimens sur 103 (73 %) de l’ouest de la baie d’Hudson ont été correctement réassignés à leur population d’origine, comparativement à 79 sur 96 (83 %) dans le cas de la population de l’est de la baie d’Hudson (de March et Postma, 2003).
Il se peut que les bélugas de l’ouest de la baie d’Hudson qui sont concentrés dans la région de Churchill forment une population distincte centrée sur le fleuve Churchill, puisqu’un fort pourcentage de ces animaux présentent un haplotype commun (de March et Postma, 2003). Comme les bélugas de l’ouest de la baie d’Hudson sont considérés comme une seule et même grande population et que le niveau d’exploitation est jugé soutenable, les chercheurs ont prélevé peu d’échantillons génétiques pour couvrir toute leur aire de répartition géographique. L’identité génétique de cette population peu étudiée intéresse cependant les gestionnaires, parce que certains individus se rendent jusque dans le détroit d’Hudson et font partie des bêtes capturées chaque année dans les collectivités du sud-est de l’île de Baffin et du détroit d’Hudson, où sont également chassés les bélugas des populations décimées du nord du Québec. Les animaux qui passent l’été au large de la côte sud-est de l’île de Baffin, mais à l’extérieur de la baie Cumberland, sont (selon la division des populations employée ici) eux aussi provisoirement considérés comme appartenant à la population de l’ouest de la baie d’Hudson (voir plus bas).
Les scientifiques étaient nombreux à penser que les bélugas qui estivent dans les eaux libres du détroit de Lancaster, du détroit de Barrows, du détroit de Peel et de la baie de Baffin, dans le haut Arctique canadien, hivernent dans les secteurs exempts de glace ou jouxtant la banquise sur la côte ouest du Groenland. Au cours du siècle dernier, les niveaux de récolte sont demeurés relativement élevés dans l’ouest du Groenland (Reeves et Mitchell, 1987).
Dans les années 1980, avant l’avènement des analyses génétiques et des études télémétriques satellitaires, les recensements aériens ont permis de confirmer la présence de bélugas près de la banquise dans des régions aussi méridionales que le 66e parallèle, dans la baie de Baffin, du côté du Groenland (Koski et Davis, 1979). Le côté ouest de la baie de Baffin, en bordure de la côte de l’île de Baffin, a toujours été caractérisé par une banquise côtière beaucoup plus extrême et par de plus fortes concentrations de glace. Des recensements aériens réalisés à la fin de l’hiver ont révélé qu’un nombre inconnu de bélugas avaient apparemment hiverné dans la partie nord de la baie de Baffin, dans la polynie des eaux du Nord (Finley et Renaud, 1980; Stirling, 1980; Richard et al., 1998a).
De 1987 à 2000, un nombre important de bélugas de l’Arctique canadien ont été marqués avec des émetteurs satellitaires, mais un seul animal sur 39 a gagné la côte ouest du Groenland (Smith et Martin, 1994; Richard et al., 1998b; Reeves et St. Aubin, 2001). En 2001, trois des cinq bélugas marqués dans la baie Creswell, dans le haut Arctique canadien, ont abouti sur la côte ouest du Groenland (Heide-Jørgensen et al., 2003). Il s’agit du même endroit d’où était parti le seul béluga marqué qui s’était rendu au Groenland (Smith et Martin, 1994). Une nouvelle analyse de l’ensemble des bélugas qui avaient conservé leur émetteur après le 1er octobre (n=26) a permis d’établir à 15 % le pourcentage des animaux qui ont gagné la côte ouest du Groenland. Cette estimation demeure fondée sur un petit échantillon, et il faut tenir compte des problèmes associés à la répartition apparemment non aléatoire des bélugas de l’ouest du Groenland pendant leur séjour dans leur aire d’estivage du haut Arctique canadien. Il devient ainsi difficile d’obtenir une idée claire de la proportion de bélugas du haut Arctique canadien qui passent l’hiver dans les eaux du Nord ou dans la région du détroit de Lancaster et de ceux qui hivernent le long de la côte ouest du Groenland (Heide-Jørgensen et al., 2003).
Les preuves génétiques donnent aussi à penser qu’au moins deux populations habitent ce vaste secteur géographique (de March et al., 2002). Un fort pourcentage des bélugas échantillonnés dans la partie ouest du détroit de Barrows et aux environs de l’île Somerset présentent l’haplotype le plus commun dans la population de l’ouest de la baie d’Hudson, lequel est moins courant chez les spécimens provenant du détroit de Peel et de la côte ouest du Groenland (de March et al., 2002). L’un des haplotypes courants chez les bélugas du détroit de Peel est également fréquent chez ceux de la côte ouest du Groenland et de la mer de Beaufort, mais il l’est moins chez ceux de la partie ouest du détroit de Barrows et de l’île Somerset. Les fréquences des allèles microsatellites varient elles aussi grandement entre ces deux régions.
Les résultats des analyses génétiques réalisées jusqu’à présent montrent des différences globales entre les bélugas du détroit de Lancaster et ceux du Groenland (de March et al., 2002). Cependant, des lots individuels d’échantillons prélevés au même endroit mais à des périodes différentes révèlent des affiliations différentes, ce qui confirme l’hypothèse selon laquelle plusieurs populations cohabiteraient dans la région pendant les mois d’été et pendant les migrations d’automne et de printemps. Il sera impossible d’obtenir une idée précise du nombre et de la répartition géographique de ces populations tant qu’un échantillonnage beaucoup plus systématique n’aura pas eu lieu.
Des Inuits de plusieurs collectivités de l’ouest du Groenland signalent la présence de bélugas particuliers dans leurs captures, et ils présument que ceux-ci viennent des eaux canadiennes (Thomsen, 1993). Ces animaux, décrits comme étant généralement plus longs et comme possédant un pédoncule caudal allongé, seraient adaptés à la nage en eaux à forts courants. Certains chasseurs font également mention de bélugas aux longues dents qu’ils appellent kigutikaat. Ils aperçoivent aussi en de rares occasions un autre type de béluga auquel ils donnent le nom de nujalik, mot qui fait allusion à la chevelure de l’animal (voir Smith et Sleno, 1986, pour une explication possible).
En résumé, les chercheurs ont réuni de nombreuses données sur la répartition (Richard et al., 1998a) et les déplacements de la population de l’est du haut Arctique et de la baie de Baffin (Heide-Jørgensen et al., 2003) ainsi que des preuves génétiques (de March et Postma, 2003) qui laissent supposer qu’elle occupe deux territoires hivernaux distincts, les eaux du Nord et la côte ouest du Groenland. On ne sait pas encore très bien si ces populations fréquentent aussi des aires d’estivage distinctes dans le haut Arctique canadien et, le cas échéant, quel est le profil de dispersion. Tout porte à croire que l’effectif des bélugas de l’ouest du Groenland a été considérablement réduit dans la dernière décennie (Heide-JØrgensen et al., 1993; Heide-JØrgensen et Aquarone, 2002) et que les niveaux de récolte actuels ne sont pas soutenables (Innes et Stewart, 2002).
Les bélugas observés dans la baie Cumberland pendant l’été sont depuis longtemps considérés comme une population distincte (Sergeant et Brodie, 1975), sans doute parce qu’ils se réunissent en grand nombre près du fond de la baie Cumberland, dans le fjord Clearwater. À cet endroit, la rivière Ranger forme un estuaire que les bélugas fréquentent selon un profil essentiellement semblable à celui de nombreux autres estuaires de l’Arctique canadien (Brodie, 1971; Sergeant et Brodie, 1975; Fraker et al., 1979; Smith et al.,1994). Il n’existe aucun autre estuaire ni aucun autre lieu de rassemblement pour les bélugas dans le sud-est de l’île de Baffin. Les bélugas de la baie Cumberland présentent une grande diversité génétique et possèdent un certain nombre d’haplotypes peu courants et des loci microsatellites qui ne se retrouvent nulle part ailleurs.
Au cours d’un réexamen des populations à des fins de gestion, Richard et Orr (1986) ont soutenu que les bélugas de la baie Cumberland appartenaient à un groupe plus dispersé de bélugas qui fréquentent aussi la région d’Iqaluit et de Kimmirut. Ils ont donné à cette population le nom collectif de population du sud-est de l’île de Baffin (Richard et Orr, 1991). Toutefois, les données récentes issues du pistage satellitaire, des analyses génétiques, de l’analyse des signatures de contaminants organochlorés et des connaissances traditionnelles autochtones (Kilabuk, 1998) semblent toutes indiquer que les bélugas de la baie Cumberland forment une population distincte. Des recensements aériens (Richard et Baratin, 2002) et des études réalisées sur des bélugas munis d’émetteurs VHF reliés à un satellite (Richard, 2002) révèlent que les individus de la baie Cumberland demeurent dans les environs immédiats, peut-être pendant tout l’hiver (P. Richard, comm. pers., MPO, Winnipeg). Les échantillons d’ADN obtenus au fil des ans depuis la fin des années 1980 permettent de conclure que les bélugas de la baie Cumberland sont distincts des autres spécimens échantillonnés ailleurs dans la région sud-est de l’île de Baffin et dans le haut Arctique (Brown-Gladden et al., 1997; de March et al., 2002).
On ignore à quelles populations appartiennent les bélugas qui occupent le reste de la région du sud-est de l’île de Baffin (voir plus bas). Les échantillons génétiques recueillis révèlent que bon nombre possèdent des haplotypes ressemblant à ceux des bélugas de l’ouest de la baie d’Hudson. Ces animaux semblent appartenir à un stock mélangé où les bélugas de l’ouest de la baie d’Hudson seraient le groupe dominant.
Il se peut que la baie Cumberland accueille plus d’une population pendant les mois d’été. Même si elles montrent des différences notables entre les bélugas qui ont été capturés à Pangnirtung, à Iqaluit et à Kimmirut, les données génétiques et les signatures de contaminants organochlorés ne contredisent pas cette hypothèse (B. de March, comm. pers., MPO, Winnipeg). Les chasseurs de la baie Cumberland reconnaissent l’existence de trois types de bélugas (Kilabuk, 1998; MPO, 2002a). Ceux qu’ils chassent à la lisière de la banquise au printemps sont plus petits, plus maigres et très blancs. Ceux qu’ils capturent en juillet et en août, dans le fjord Clearwater, sont plus gros et ont une coloration jaune qui témoigne de la mue de l’épiderme. Ceux qu’ils chassent à l’extérieur du fjord Clearwater, du côté ouest de la baie Cumberland, sont plus petits et plus maigres; leur épiderme (maayak ou muktuk) est plus épais et a un goût plus fort.
Dans l’ensemble, les données récentes sur le profil génétique, la répartition et l’abondance viennent corroborer l’idée que les bélugas de la baie Cumberland forment une population distincte.
Une importante population de bélugas passe le début de l’été dans le sud-est de la mer de Beaufort et fréquente l’estuaire du Mackenzie (Fraker et al., 1979; Harwood et al., 1996; MPO, 2000). Fraker (1980), sans posséder de preuve directe, considérait cette population comme distincte en raison de son profil migratoire connu et de l’absence de concentration voisine de bélugas durant l’été le long de la côte de l’Alaska. Des recensements aériens plus récents (Harwood et al., 1996) et des études de marquage (Richard et al., 2001b) sont venus donner du poids à ses affirmations.
Des analyses génétiques ont permis de confirmer que les bélugas de la mer de Beaufort sont nettement distincts de ceux des autres populations de l’espèce au Canada (Brown-Gladden et al., 1997) et de ceux des populations voisines de l’Alaska, avec lesquels ils pourraient partager des lieux d’hivernage (Brown-Gladden et al., 1997; O’Corry-Crowe et al., 1997; Brown-Gladden et al., 1999). Cette séparation est basée principalement sur les haplotypes d’ADN mitochondrial hérités de la mère et, dans une certaine mesure, sur les données relatives aux microsatellites. Ainsi, il se peut que ces deux populations s’accouplent entre elles en hiver, mais elles se séparent pour gagner des zones d’estivage différentes.
La division des populations présentée ci-dessus comporte trois zones grises. Les voici :
- Sud-est de l’île de Baffin. L’identité des animaux observés au large de la partie sud-est de l’île de Baffin, mais à l’extérieur de la baie Cumberland, demeure incertaine. Les données les plus récentes indiquent que ces bélugas n’appartiennent pas à la population de la baie Cumberland. Les chasseurs inuits d’Iqaluit et de Kimmirut, sur la côte sud-est de l’île de Baffin, font état de captures de tailles et de formes différentes et croient chasser des spécimens appartenant à plusieurs populations différentes (Kilabuk, 1998). Iqaluit et Kimmirut se trouvent près des aires d’hivernage présumées des populations de la baie d’Hudson, et les chasseurs de ces deux villages récoltent des animaux qui semblent faire partie de ces populations. Certaines données sur les caractéristiques génétiques et les contaminants organochlorés (de March et al., sous presse) recueillies sur des bélugas du sud-est de l’île de Baffin vont dans ce sens. Comme les échantillons individuels ou groupés qui ont été prélevés pendant la migration printanière ou automnale représentent sans doute plusieurs populations de bélugas, les régions telles que le sud-est de l’île de Baffin, qui sont adjacentes à un territoire d’hivernage partagé par plusieurs populations, présentent un défi particulier sur le plan de l’identification et de la gestion des populations. Les animaux qui fréquentent le sud-est de l’île de Baffin en été pourraient avoir les plus grandes affinités avec les populations de la baie d’Hudson.
- Sud de la baie d’Hudson. Il existe peu de données sur les animaux qui passent l’été dans la baie James et dans les eaux voisines, au large du littoral de l’Ontario. Les données obtenues (voir plus haut) donnent cependant à penser que ces bélugas ont un lien de parenté plus étroit avec la population de l’ouest de la baie d’Hudson qu’avec celle de l’est de la baie d’Hudson.
- Haut Arctique. Les chercheurs possèdent un nombre croissant de preuves attestant que la population de l’est du haut Arctique et de la baie de Baffin a une importante sous-structure (voir plus haut). Cependant, les données recueillies à ce jour permettent de conclure qu’il n’est probablement pas justifié de subdiviser cette population, selon les lignes directrices du COSEPAC.
Détails de la page
- Date de modification :