
Paruline du Canada (Cardellina canadensis) : programme de rétablissement 2016
Loi sur les espèces en péril
Série de Programmes de rétablissement
Paruline du Canada

2016
Table des matières
- Préface
- Remerciements
- Sommaire
- Résumé du caractère réalisable du rétablissement
- 1. Évaluation de l'espèce par le Comité sur la situation des espèces en péril au Canada (COSEPAC)
- 2. Information sur la situation de l'espèce
- 3. Information sur l'espèce
- 4. Menaces
- 5. Objectifs de population et de répartition
- 6. Stratégies et approches générales recommandés pour l'atteinte des objectifs
- 7. Habitat essentiel
- 8. Mesure des progrès
- 9. Énoncé sur les plans d'action
- 10. Références
Information sur le document
Programme de rétablissement de la Paruline du Canada (Cardellina canadensis) au Canada - 2016

Référence recommandée :
Environnement Canada. 2016. Programme de rétablissement de la Paruline du Canada (Cardellina canadensis) au Canada, Série de Programmes de rétablissement de la Loi sur les espèces en péril, Environnement Canada, Ottawa, vii + 62 p.
Pour télécharger le présent programme de rétablissement ou pour obtenir un complément d'information sur les espèces en péril, incluant les rapports de situation du COSEPAC, les descriptions de la résidence, les plans d'action et d'autres documents connexes sur le rétablissement, veuillez consulter le Registre public des espèces en péril.
Illustration de la couverture : Paruline du Canada photographiée par Henry Skrzypkowski © Frontier Digital Arts.
Also available in English under the title
"Recovery Strategy for the Canada Warbler (Cardellina canadensis) in Canada"
Le contenu du présent document (à l'exception des illustrations) peut être utilisé sans permission, mais en prenant soin d'indiquer la source.
Préface
En vertu de l'Accord pour la protection des espèces en péril (1996), les gouvernements fédéral, provinciaux et territoriaux signataires ont convenu d'établir une législation et des programmes complémentaires qui assureront la protection efficace des espèces en péril partout au Canada. En vertu de la Loi sur les espèces en péril (L.C. 2002, ch. 29) (LEP), les ministres fédéraux compétents sont responsables de l'élaboration des programmes de rétablissement pour les espèces inscrites comme étant disparues du pays, en voie de disparition ou menacées et sont tenus de rendre compte des progrès réalisés dans les cinq ans suivant la publication du document final dans le Registre public des espèces en péril.
La ministre de l'Environnement et ministre responsable de l'Agence Parcs Canada est la ministre compétente en vertu de la LEP de la Paruline du Canada et a préparé ce programme de rétablissement, conformément à l'article 37 de la LEP. Dans la mesure du possible, il a été préparé en collaboration avec les gouvernements de la Colombie Britannique, de l'Alberta, de la Saskatchewan, du Manitoba, de l'Ontario, du Québec (ministère des Forêts, de la Faune et des Parcs), du Nouveau Brunswick, de l'Île du-Prince-Édouard et de la Nouvelle-Écosse, les gouvernements du Yukon et des Territoires du Nord-Ouest, et d'autres parties intéressées, en vertu du paragraphe 39(1) de la LEP.
La réussite du rétablissement de l'espèce dépendra de l'engagement et de la collaboration d'un grand nombre de parties concernées qui participeront à la mise en œuvre des directives formulées dans le présent programme. Cette réussite ne pourra reposer seulement sur Environnement Canada et l'Agence Parcs Canada, ou sur toute autre autorité responsable. Tous les Canadiens et les Canadiennes sont invités à appuyer ce programme et à contribuer à sa mise en œuvre pour le bien de la Paruline du Canada et de l'ensemble de la société canadienne.
Le présent programme de rétablissement sera suivi d'un ou de plusieurs plans d'action qui présenteront de l'information sur les mesures de rétablissement qui doivent être prises par Environnement Canada, l'Agence Parc Canada, et d'autres autorités responsables et/ou organisations participant à la conservation de l'espèce. La mise en œuvre du présent programme est assujettie aux crédits, aux priorités et aux contraintes budgétaires des autorités responsables et organisations participantes.
Le programme de rétablissement établit l'orientation stratégique visant à arrêter ou à renverser le déclin de l'espèce, incluant la désignation de l'habitat essentiel dans la mesure du possible. Il fournit à la population canadienne de l'information pour aider à la prise de mesures visant la conservation de l'espèce. Lorsqu'un programme de rétablissement désigne de l'habitat essentiel, il peut y avoir des incidences réglementaires futures, selon l'endroit où se trouve l'habitat essentiel désigné. La LEP exige que l'habitat essentiel désigné se trouvant à l'intérieur d'aires protégées fédérales soit décrit dans la Gazette du Canada, après quoi les interdictions relatives à la destruction de cet habitat seront appliquées. En ce qui concerne l'habitat essentiel situé sur le territoire domanial à l'extérieur des aires protégées fédérales, la ministre de l'Environnement doit présenter un énoncé sur la protection juridique existante ou prendre un arrêté de manière à ce que les interdictions relatives à la destruction de l'habitat essentiel soient appliquées. En ce qui concerne l'habitat essentiel se trouvant sur le territoire non domanial, si la ministre de l'Environnement estime qu'une partie de l'habitat essentiel n'est pas protégée par les dispositions de la LEP, par les mesures prises aux termes de cette dernière ou par toute autre loi fédérale, et que cette partie de l'habitat essentiel n'est pas protégée efficacement par les lois provinciales ou territoriales, elle doit, comme le prévoit la LEP, recommander au gouverneur en conseil de prendre un décret visant à étendre l'interdiction de détruire à cette partie de l'habitat essentiel. La décision de protéger l'habitat essentiel se trouvant sur le territoire non domanial et n'étant pas autrement protégé demeure à la discrétion du gouverneur en conseil.
Remerciements
Le présent programme de rétablissement a été établi par Krista Baker, Julie McKnight, Andrew Horn et Peter Thomas (Service canadien de la faune d'Environnement Canada (SCF – EC), Région de l'Atlantique), d'après une première ébauche rédigée par Kimberley Hair et Madison Wikston (SCF – EC, Région de la capitale nationale). De nombreuses personnes ont revu les ébauches et fourni de précieux commentaires : Manon Dubé et Adam Smith (SCF – EC, Région de la capitale nationale), Andrew Boyne, David Andrews, Samara Eaton et Becky Whitham (SCF – EC, Région de l'Atlantique), Gilles Falardeau, Vincent Carignan, Bruno Drolet, Josée Tardif, Renée Langevin et Mireille Poulin (SCF – EC, Région du Québec), Mark Bidwell, Lisa Mahon, Jeff Ball, Samuel Haché et Steven Van Wilgenburg (SCF – EC, Région des Prairies et du Nord), Pam Sinclair, Craig Machtans, Krista De Groot et Wendy Easton (SCF – EC, Région du Pacifique et du Yukon), Kathy St. Laurent, Rich Russell, Kevin Hannah, Russ Weeber, Madeline Austen, Lesley Dunn, Elizabeth Rezek, Christian Friis et Krista Holmes (SCF – EC, Région de l'Ontario), François Fournier, Scott Wilson, Greg Mitchell et Junior Tremblay (Sciences et technologie – EC).
Nous remercions tous les autres qui ont fourni des conseils et des commentaires ayant servi à étayer l'élaboration de ce programme de rétablissement, dont des Autochtones et des organisations autochtones, des gouvernements provinciaux et territoriaux, d'autres ministères fédéraux (dont le ministère de la Défense nationale), des propriétaires fonciers, des citoyens et divers intervenants.
Sommaire
La Paruline du Canada (Cardellina canadensis) est un petit oiseau chanteur forestier. Elle se reproduit généralement dans des forêts mixtes ou des forêts de feuillus à sous-étage dense et complexe, dans lesquelles la composition en espèces d'arbres et l'importance de la topographie et des aires humides présentent une variation géographique à l'échelle de l'aire de répartition canadienne de l'espèce.
L'espèce a été désignée « menacée » par le Comité sur la situation des espèces en péril au Canada (COSEPAC) en 2008. Depuis 2010, elle est inscrite comme telle à l'annexe 1 de la Loi sur les espèces en péril (LEP). La population canadienne, qui occupe 82 % de l'aire de répartition mondiale de l'espèce, compte environ 3 millions d'individus, soit 75 % de la population mondiale. L'espèce niche dans l'ensemble des provinces et des territoires du Canada sauf au Nunavut et à Terre-Neuve-et-Labrador.
Les principales menaces pesant sur la Paruline du Canada comprennent la conversion des terres de l'habitat de reproduction et de l'habitat autre que l'habitat de reproduction, l'exploitation forestière et la sylviculture, l'enlèvement d'arbustes, l'exploration et l'exploitation de ressources énergétiques et minières, le broutage excessif, la réduction de la disponibilité d'insectes proies, et les collisions contre les fenêtres. L'importance de chacune de ces menaces varie dans l'aire de répartition de l'espèce.
Le rétablissement de la Paruline du Canada est jugé réalisable, mais le potentiel de rétablissement demeure incertain du fait de plusieurs facteurs inconnus. Malgré les incertitudes, et conformément au principe de précaution, un programme de rétablissement a été élaboré aux termes du paragraphe 41(1) de la LEP.
Le présent programme de rétablissement fixe des objectifs à court et à long terme pour la Paruline du Canada. L'objectif de population à court terme est d'arrêter le déclin national d'ici 2025 (c'est à dire dans les 10 ans suivant la publication du présent programme de rétablissement dans le Registre public des espèces en péril), en veillant à ce que la population ne diminue pas de plus de 10 % durant cette période. L'objectif de population à long terme (après 2025) est d'assurer une tendance démographique positive sur 10 ans à l'échelle du pays. L'objectif de répartition est de maintenir la zone d'occurrence actuelle (zone englobant la répartition géographique de l'ensemble des populations connues) au Canada. Les stratégies et approches générales visant à atteindre ces objectifs sont décrites dans le présent programme de rétablissement.
Pour l'instant, les renseignements dont on dispose sont insuffisants pour désigner l'habitat qui est nécessaire à la survie ou au rétablissement de la Paruline du Canada. Un calendrier des études visant à obtenir les renseignements nécessaires à la désignation de l'habitat essentiel est présenté.
Un ou plusieurs plans d'action seront publiés dans le Registre public des espèces en péril dans les cinq ans suivant la publication du présent programme de rétablissement.
Résumé du caractère réalisable du rétablissement
D'après les quatre critères suivants appliqués par Environnement Canada, le caractère réalisable du rétablissement de la Paruline du Canada comporte des inconnues. Néanmoins, conformément au principe de précaution, le présent programme de rétablissement a été préparé en vertu du paragraphe 41(1) de la LEP, comme il convient de faire lorsque le rétablissement est jugé réalisable. Ce programme de rétablissement prend en considération les inconnues entourant le caractaire réalisable du rétablissement de l'espèce.
- 1. Des individus de l'espèce sauvage capable de se reproduire sont disponibles maintenant ou le seront dans un avenir prévisible pour maintenir la population ou augmenter son abondance.
Oui. L'espèce occupe encore l'ensemble de son aire de répartition et des individus nicheurs sont actuellement présents un peu partout au Canada, ainsi qu'aux États Unis. La population canadienne est estimée à 3 000 000 d'individus. On pense qu'il existe aujourd'hui suffisamment d'individus pour maintenir la population canadienne ou accroître son effectif moyennant la mise en œuvre de mesures de conservation appropriées. - De l'habitat convenable suffisant est disponible pour soutenir l'espèce ou pourrait être rendu disponible par des activités de gestion ou de remise en état de l'habitat.
On ne sait pas. L'habitat convenable à la reproduction est probablement assez abondant pour assurer le maintien de l'espèce, et on pourrait en créer par des mesures d'aménagement (p. ex. pratiques forestières qui préservent la strate arbustive, conservent les arbres servant de perchoirs, et font que les tapis forestiers demeurent humides, ou conservent au voisinage de corridors riverains de grandes étendues de vieille forêt dominée par les feuillus à strate arbustive dense) ou de remise en état. Les caractéristiques de l'habitat convenable à l'échelle du paysage ne sont pas suffisamment bien connues pour qu'on puisse établir de façon exhaustive les besoins de l'espèce en matière d'habitat aux fins de son rétablissement.
La Paruline du Canada peut tolérer un certain degré de perturbation de son habitat (p. ex. cultures de cardamome sous ombrage) dans les zones autres que les zones de reproduction, mais, en général, son habitat d'hivernage en Amérique du Sud va en diminuant. On ne sait donc pas s'il reste suffisamment d'habitat convenable pour l'espèce dans son aire d'hivernage. - Les principales menaces qui pèsent sur l'espèce ou sur son habitat (y compris les menaces qui proviennent de l'extérieur du Canada) peuvent être évitées ou atténuées.
On ne sait pas. On pense que les principales menaces pesant sur l'habitat de l'espèce dans ses lieux de reproduction au Canada (p. ex. l'exploitation forestière et la sylviculture, la conversion des terres, l'enlèvement d'arbustes, et le broutage excessif) peuvent être atténuées ou évitées grâce à des mesures de conservation ciblées reposant sur des activités de recherche et d'intendance appropriées.
La dégradation et la perte d'habitat d'hivernage pourraient constituer une menace importante pour l'espèce. Cependant, on ne sait pas bien s'il existe une relation directe de cause à effet entre les baisses d'effectifs et la disponibilité d'habitat d'hivernage. Même si cette relation en venait à être clairement établie, on ne sait pas très bien quels moyens pourraient être employés pour protéger ou remettre en état l'habitat d'hivernage de l'espèce. Néanmoins, de nombreux programmes et organisations (p. ex. le programme Southern Wings, le programme des Zones importantes pour la conservation des oiseaux) s'efforcent actuellement de conserver l'habitat d'hivernage des oiseaux migrateurs néotropicaux.
Des menaces possiblement importantes, autres que celles liées à la disponibilité d'habitat, ont été repérées, mais la plupart n'ont pas été étudiées ou vérifiées, comme la réduction de la disponibilité d'insectes proies et les collisions durant les migrations. Ces menaces pourraient probablement être atténuées au moyen de mesures de conservation ciblées. - Il existe des techniques de rétablissement permettant d'atteindre les objectifs en matière de population et de répartition, ou de telles techniques pourraient être élaborées dans un délai raisonnable.
On ne sait pas. On pense qu'une des principales approches pour le rétablissement consistera à maintenir l'habitat de reproduction et l'habitat autre que l'habitat de reproduction. La gestion et l'intendance de l'habitat pourraient être efficaces pour cette espèce. Il pourrait ne pas y avoir de problème de disponibilité de l'habitat convenable dans les lieux de reproduction de l'espèce. Il faudra effectuer des travaux de recherche pour déterminer les éléments de l'habitat essentiel de la Paruline du Canada et aider à concevoir des pratiques d'utilisation des terres et de gestion de l'habitat qui profiteront à l'espèce dans les zones de reproduction et zones autres que les zones de reproduction. Des mesures de conservation ciblées pourraient atténuer la menace que constituent les collisions contre les fenêtres durant les migrations, dans un délai raisonnable.
Un défi de taille sera de réaliser les recherches nécessaires sur l'importance de la perte d'habitat d'hivernage, et de travailler à protéger l'habitat convenable qui existe. Des programmes et organisations en place pourraient être utilisés (dans la mesure du possible) pour faire en sorte que les mesures susmentionnées soient appliquées dans un délai raisonnable.
1. Évaluation de l'espèce par le COSEPACNote de bas de page1
Sommaire d'évaluation
- Date de l'évaluation :
- Avril 2008
- Nom commun (population) :
- Paruline du Canada
- Nom scientifique :
- Cardellina canadensis Note de bas de page2
- Statut selon le COSEPAC :
- Espèce menacée
- Justification de la désignation :
- La majeure partie (80 %) de l'aire de reproduction de l'espèce se trouve au Canada. Quoique les tendances régionales puissent varier, dans l'ensemble l'espèce a connu un important déclin à long terme. Ce déclin est particulièrement évident dans le cas de l'aire de répartition canadienne de l'espèce, et il n'y a aucune indication que la tendance se renversera. Les raisons de ce déclin sont peu connues, mais la perte de forêt primaire dans l'aire d'hivernage en Amérique du Sud est une cause potentielle.
- Présence au Canada :
- Yukon, Territoires du Nord-Ouest, Colombie-Britannique, Alberta, Saskatchewan, Manitoba, Ontario, Québec, Nouveau-Brunswick, Île-du-Prince-Édouard, Nouvelle-Écosse
- Historique du statut établi selon le COSEPAC:
- Espèce désignée « menacée » en avril 2008.
2. Information sur la situation de l'espèce
Le Canada comprend environ 82 % de l'aire de reproduction mondiale de la Paruline du Canada (Cardellina canadensis) (Partners in Flight Science Committee, 2012) et accueille quelque 75 % des individus nicheurs de l'espèce (Partners in Flight Science Committee, 2013). L'espèce a été inscrite comme espèce menacée à l'annexe 1 de la Loi sur les espèces en péril (L.C. 2002, chap. 29) en 2010. Le tableau 1 présente les cotes de conservation nationales qui lui sont attribuées aux États-Unis et au Canada, ainsi que les cotes infranationales au Canada. Selon les lois provinciales qui protègent les espèces en péril, la Paruline du Canada est une espèce préoccupante en Ontario (L.O. 2007, chap. 6), menacée au Nouveau-Brunswick (L.N.-B. 2012, chap. 6) et au Manitoba (C.P.L.M., chap. E111), et en voie de disparition en Nouvelle-Écosse (S.N.S. 1998, chap. 11). Au Québec, l'espèce figure sur la liste des espèces fauniques susceptibles d'être désignées menacées ou vulnérables, établie en vertu de la Loi sur les espèces menacées ou vulnérables (RLRQ, chap. E-12.01). En Colombie-Britannique, la Paruline du Canada figure sur la liste bleue (liste des espèces préoccupantes), ce qui ne lui confère toutefois aucune protection juridique. L'espèce ne figure actuellement sur aucune liste d'espèces en péril au Yukon, dans les Territoires du Nord-Ouest, en Alberta, en Saskatchewan et à l'Île-du-Prince-Édouard. Elle figure sur la liste de surveillance États-Unis-Canada, la liste trinationale des espèces fortement préoccupantes (Species of High Tri-National Concern List) et la liste d'intendance États-Unis-Canada de Partenaires d'envol (Partners in Flight Science Committee, 2012), parce qu'elle présente des tendances à la baisse et qu'elle est exposée à des menaces importantes. La liste de surveillance renferme les espèces les plus préoccupantes à l'échelle continentale. La cote de conservation mondiale attribuée à la Paruline du Canada par NatureServe est G5 – non en péril (NatureServe, 2013). Le tableau 1 présente d'autres cotes NatureServe pour l'espèce.
| Cote mondiale (G)Note de bas de pagea | Cote nationales (N)Note de bas de pageb | Cote infranationale (S)Note de bas de pagec |
|---|---|---|
| G5 | Canada N5B (12 févr. 2012) États-Unis N5B (19 mars 1997) |
Alberta (S3S4) Colombie-Britannique (S3S4B) Manitoba (S4B) Nouveau-Brunswick (S3S4B) Île de Terre-Neuve (S1) Territoires du Nord-Ouest (SUB) Nouvelle-Écosse (S3B) Ontario (S4B) Île-du-Prince-Édouard (S3B) Québec (S3S4) Saskatchewan (S5B) Yukon (S1B) |
3. Information sur l'espèce
3.1 Description de l'espèce
La Paruline du Canada, oiseau chanteur (passereau) de petite taille, a le dos gris-bleu et les parties inférieures jaunes. Le mâle a les yeux nettement cerclés de jaune et présente un collier formé de raies noires contrastantes sur la poitrine. Le plumage de la femelle présente des caractéristiques similaires, mais est plus terne, et celui des jeunes est plus terne encore. Les Parulines du Canada sont longues d'entre 12 et 14 cm, et pèsent en général de 10 à 12 grammes, en partie selon leurs gains de poids saisonniers (Reitsma et al., 2010). Le chant du mâle, court et puissant, consiste généralement en un « tchip » sec suivi d'un enchaînement rapide et irrégulier de riches notes modulées.
3.2 Population et répartition
La Paruline du Canada niche dans les régions de la forêt boréale méridionale et de la forêt mixte, incluant la forêt des Grands Lacs et du Saint-Laurent et la partie nord des plaines à forêt mixte, jusque dans le sud des plaines hudsoniennes. Elle est présente dans l'extrême sud-est du Yukon, le nord-est de la Colombie-Britannique, le sud-ouest des Territoires du Nord-Ouest, le nord de l'Alberta, le centre de la Saskatchewan et du Manitoba, le centre et le sud de l'Ontario et du Québec, et les Maritimes (figure 1). Aux États Unis, l'aire de reproduction de l'espèce s'étend vers le sud jusque dans le nord est du Minnesota, le nord du Wisconsin, le Michigan, l'Ohio, et, de façon discontinue, dans les Appalaches jusque dans l'extrême nord de la Géorgie (figure 1). Dans la partie ouest de son aire, elle peut nicher occasionnellement vers le sud jusque dans le Dakota du Nord et l'Iowa (Reitsma et al., 2010). L'aire de l'espèce pourrait s'être rétrécie dans le sud ouest de l'Ontario, la péninsule inférieure du Michigan et l'est de la Pennsylvanie, et s'être étendue en Colombie-Britannique, dans le centre-sud de l'Ontario et dans l'État de New York, par suite de changements de la disponibilité d'habitat au cours du siècle dernier (Reitsma et al., 2010).
Généralement, comparativement aux autres parulines, la Paruline du Canada arrive tard dans ses lieux de reproduction, entreprend tôt sa migration automnale et migre rapidement sur une courte période (Reitsma et al., 2010). Sanders et Mennill (2014a) l'ont enregistré (détection des cris de vol de nuit) traversant le lac Érié. La Paruline du Canada est un migrateur néotropical qui hiverne dans l'extrême nord du Brésil, au Venezuela, au Panama, dans le nord de la Colombie, et sur le versant oriental des Andes en Équateur et dans le nord du Pérou (figure 1).
Figure 1. Aires de reproduction, de migration et d'hivernage de la Paruline du Canada (carte de BirdLife International et NatureServe (2013) modifiée à partir de données de Haché et al. (2014) et de eBird (2014)).
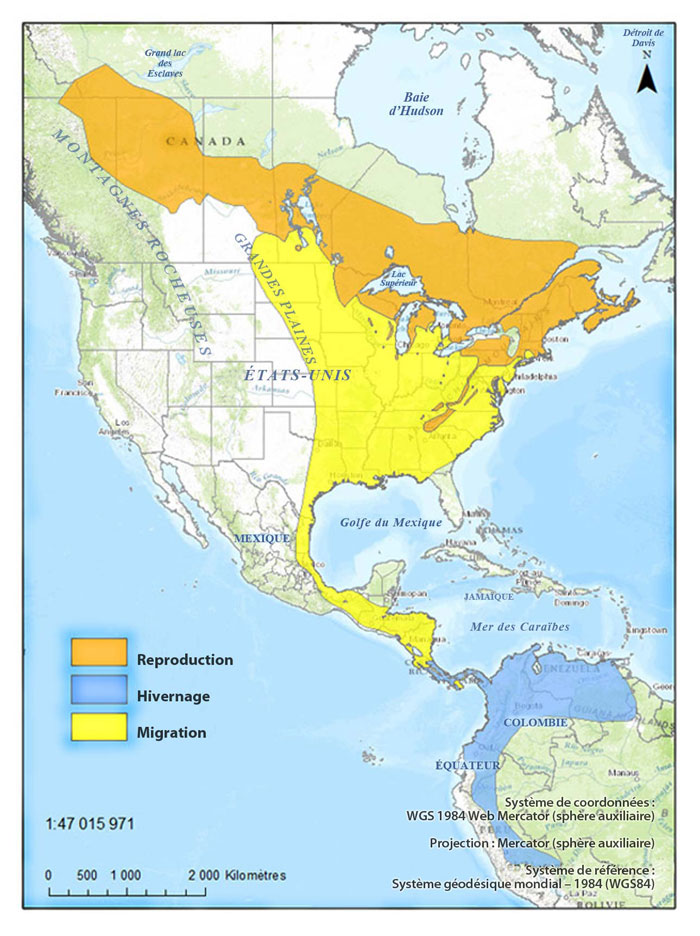
Carte : © Environnement Canada
Description longue pour la Figure 1
La figure 1 illustre l'aire de répartition de l'espèce en Amérique du nord, en Amérique centrale et en Amérique du sud, selon le type d'aire. L'aire de reproduction s'étend du sud des Territoires du Nord Ouest au Canada, vers la cote nord est des États Unis. L'aire de migration se situe principalement dans l'est des États Unis; elle s'étend du Dakota du Nord vers le Texas et vers la cote est. Elle progresse ensuite vers le sud le long de la cote est du Mexique jusqu'au Costa Rica. L'aire d'hivernage commence au Costa Rica et s'étend jusqu'au nord de l'Amérique du sud, du Venezuela au Pérou.
Le rapport de situation du COSEPAC a repris les données du document de Partenaires d'envol de 2004 pour établir une estimation de la population de 1,4 million d'individus (Rich et al., 2004). Plus récemment, la base de données sur les estimations de populations de Partenaires d'envol (Partners in Flight Population Estimates) a été mise à jour et constitue aujourd'hui la source d'information la plus complète sur les oiseaux terrestres d'Amérique du Nord. La nouvelle estimation de la population de Parulines du Canada présentée dans cette base de données est de 4 millions d'individus, dont 3 millions au Canada (Partners in Flight Science Committee, 2013). La qualité des données ayant servi à l'estimation de Partenaires d'envol est cotée « beige », deuxième meilleure cote, après « vert ». Cette cote « beige » a trait au niveau de variance associé au dénombrement moyen, et indique que les limites de confiance à 95 % de part et d'autre du dénombrement moyen pour la région considérée se situent à moins de 50 % de cette valeur moyenne (Blancher et al., 2007). Ce changement de l'estimation de la population ne représente pas un accroissement réel de la population, mais résulte plutôt de l'application de nouvelles techniques analytiques. Il découle en grande partie d'un ajustement de la distance de détection utilisée pour estimer la densité. Vu l'incertitude dans l'estimation des effectifs absolus, nous avons centré notre évaluation de l'état de la population sur la tendance de son effectif dans le présent programme de rétablissement.
Les dernières analyses des données du Relevé des oiseaux nicheurs (BBS, pour Breeding Bird Survey) réalisées par Environnement Canada révèlent pour la population canadienne une tendance annuelle de -2,9 % entre 1970 et 2012, les limites inférieure et supérieure de l'intervalle de crédibilité à 95 % pour cette valeur étant de -4,07 et -1,78, respectivement (Environnement Canada, 2014c) (figure 2). Ce taux annuel de diminution correspond à une baisse de la population d'environ 71 % de 1970 à 2012 (Adam Smith, comm. pers.). Cependant, l'interprétation des données du BBS présente des difficultés fondamentales à cause de problèmes de couverture et de biais (Machtans et al., 2014). Le BBS n'offre pas un échantillonnage aléatoire couvrant l'entièreté de l'aire de reproduction de l'espèce : la majorité des parcours du BBS sont de façon générale situés dans les régions méridionales du Canada qui sont perturbées par l'activité humaine, ce qui peut biaiser les estimations des effectifs des espèces dont l'aire de répartition s'étend dans le nord, comme la Paruline du Canada (Machtans et al., 2014). En outre, les données du BBS sous-estiment généralement les densités de Parulines du Canada, parce que les observations sont faites en bordure des routes (Haché et al., 2014). Pour toutes ces raisons, l'estimation de la population et de ses tendances à l'échelle de l'aire de l'espèce est frappée d'incertitude.
Les résultats du BBS pour le sud ouest des Territoires du Nord Ouest montrent que l'effectif de Parulines du Canada y a diminué annuellement de 0,85 % entre 1998 et 2014 (S. Wilson, comm. pers.). En Ontario, selon le Programme de surveillance des oiseaux forestiers, la tendance annuelle de l'effectif dans les régions de conservation des oiseaux 12 et 13 est de -6,3 % (A. Smith, comm. pers.). Il se pourrait que l'Ontario abrite environ 50 % de la population canadienne de Parulines du Canada, et selon l'Atlas des oiseaux nicheurs de l'Ontario, la tendance annuelle de l'effectif de la province a été de 0,8 % (intervalle de confiance : de -2 à 0,3) entre la première période de l'atlas (1981-1985) et la deuxième (2001-2005) (Environnement Canada, 2011b). Par ailleurs, l'Atlas des oiseaux nicheurs des Maritimes a révélé des baisses significatives de la probabilité d'observation de la paruline dans presque toute la région entre sa première période (1986-1990) et sa deuxième (2006-2010) (Atlas des oiseaux nicheurs des Maritimes, données inédites). L'atlas des oiseaux nicheurs de l'Alberta a montré que la répartition de l'espèce n'a pas changé et que son effectif a augmenté dans la province entre 1987-1992 et 2000-2005 (Semenchuk, 2007).
Figure 2. Estimations des tendances à long terme pour la Paruline du Canada au Canada (1970 2012) (Environment Canada, 2014). Indice = estimation du nombre moyen d'oiseaux qui seraient dénombrés par un observateur moyen sur des parcours du Relevé des oiseaux nicheurs au cours d'une année donnée
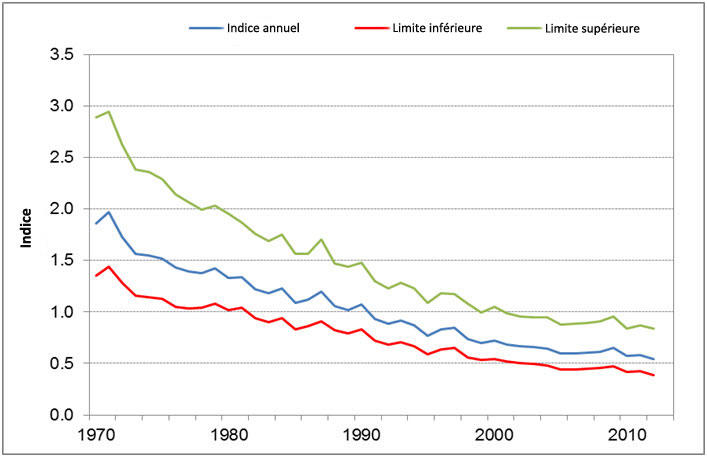
Carte : © Environnement Canada 2014
Description longue pour la Figure 2
La figure 2 montre trois courbes dans un graphique dans lequel l'axe horizontal représente les années et l'axe vertical un indice d'abondance allant de 0 à 3,5 suivant une échelle divisée en intervalles de 0,5. La limite inférieure de la tendance commence à 1,3 en 1970, monte légèrement jusqu'à 1,4 en 1972, puis descend lentement jusqu'à 0,4 en 2010. L'indice annuel commence à 1,8, monte légèrement jusqu'à 1,9, en 1972, puis descend lentement jusqu'à 0,5 en 2010. La troisième courbe, soit la limite supérieure, commence à 2,8, monte légèrement jusqu'à 2,9 en 1972, puis descend d'abord rapidement et ensuite lentement jusqu'à 0,8 en 2010.
3.3 Besoins de la Paruline du Canada
La connaissance actuelle des besoins écologiques de la Paruline du Canada est biaisée par le choix des sites d'étude (et les résultats associés), qui est tributaire de l'accessibilité de ces derniers. De plus, les besoins apparents de l'espèce en matière d'habitat pourraient être en partie faussés par le fait que la disponibilité actuelle d'habitat présenterait des limitations par rapport au passé.
Reproduction et alimentation
Type de forêt
La Paruline du Canada se reproduit dans divers milieux qui diffèrent selon l'endroit de son aire de répartition, mais presque toujours dans des forêts humides présentant une strate dense d'arbustes feuillus, un sous-étage complexe et des arbres pouvant servir de perchoirs. Néanmoins, ses milieux de prédilection peuvent présenter des caractéristiques relativement particulières aux échelles régionale et locale.
Les préférences de l'espèce en matière d'habitat et les descriptions de son habitat établies à ce jour comprennent notamment les suivantes :
- Nord-est de la Colombie-Britannique : La Paruline du Canada occupe habituellement les sites où le peuplier faux-tremble représente la principale espèce du couvert forestier (en moyenne 62 %). Le couvert arbustif était en moyenne de 79 %, et la hauteur de la végétation arbustive variait en moyenne de 2,5 à 3,5 m (Campbell et al., 2007).
- Alberta – L'espèce est le plus commune dans les vieilles tremblaies (et forêts mixtes dans une moindre mesure) (de plus de 120 ans d'âge) (Schieck et al., 1995; Hobson et Schieck, 1999; Schieck et Song, 2006; Ball et al., 2013). Des Parulines du Canada ont aussi été observées en plus faibles densités dans des milieux de début de succession (p. ex. des peuplements mixtes de 22 à 28 ans après récolte) où une moyenne de 6 % des arbres qui formaient l'étage supérieur avant récolte (Hobson et Schieck, 1999) ou de grandes parcelles boisées avaient été conservées (Ball et al., 2013), mais, notablement, aucune n'a été observée dans de jeunes peuplements (23-26 ans) dans le centre-est de l'Alberta (Schieck et al., 1995).
- Saskatchewan – L'espèce a été observée dans des tremblaies jeunes pures, matures et vieilles (Hobson et Bayne, 2000a; Hobson et Bayne, 2000b), mais surtout dans des tremblaies matures (Hobson et Bayne, 2000a). Une étude a montré que l'espèce était significativement plus commune en forêt continue que dans des fragments de forêt entourés de terres agricoles (Hobson et Bayne, 2000c).
- Ontario – L'espèce est présente dans la forêt mixte boréale. Des densités particulièrement élevées ont été observées dans une forêt sise en terrain élevé qui avait été exploitée environ six ans auparavant, présentant un couvert fermé à 72 % et une importante couverture d'arbustes de haute taille et de plantes herbacées (Welsh et Fillman, 1980).
- Québec – L'espèce est le plus commune dans des forêts mixtes boréales caractérisées par des perturbations naturelles, mais est aussi présente (à moins d'endroits) dans des forêts mixtes boréales modifiées par des aménagements préindustriels ou l'exploitation forestière industrielle (Drapeau et al., 2000).
- Maritimes – L'espèce a été observée dans des cédrières marécageuses matures et autres milieux humides, dans des forêts mixtes complexes, matures ou en régénération, dans des secteurs de coupe partielle et dans des zones arbustives (Atlas des oiseaux nicheurs des Maritimes, données inédites).
- Nouvelle-Écosse – Sites humides où poussent l’osmonde cannelle, l’aulne rugueux ou d’autres arbustes décidus. Les oiseaux sont associés avec des sols inégaux recouverts de sphaigne (A. Westwood, comm. pers.).
- New Hampshire – L'espèce a été observée dans un marécage à érable rouge dominé par l'érable rouge, le sapin baumier et l'épinette rouge, présentant une couverture au sol complexe, un sous-étage dense et des eaux stagnantes et courantes. Les préférences en matière d’habitat incluent également la forêt mixte en régénération de 6 à 20 ans après coupe renfermant des arbres résiduels et une végétation de sous-étage de moins de 6 m de haut (Hallworth et al., 2008a).
Dans une étude couvrant l'ensemble de l'aire de reproduction canadienne de la Paruline du Canada, Haché et al. (2014) ont observé que les densités de l'espèce étaient généralement plus élevées dans les peuplements mixtes et les peuplements de feuillus renfermant de grands arbres que dans d'autres types d'habitat (Haché et al., 2014). Ces auteurs ont aussi observé que les densités étaient négativement corrélées avec la proportion de terres aménagées à des fins agricoles ou autres dans une zone de 16 km² (Haché et al., 2014).
Enfin, divers autres milieux abritant des Parulines du Canada ont aussi été décrits de façon générale : forêts marécageuses, fourrés d'arbustes marécageux, bois riverains, forêts humides, ravins broussailleux, forêts de feuillus nordiques, forêts matures dont le couvert présente des ouvertures, muskeg ouvert ou arboré, etc. (Hallworth et al., 2008b; Chace et al., 2009; Reitsma et al., 2010).
Échelle locale
Durant la reproduction, la Paruline du Canada défend un territoire dont la taille est de 1 ha en moyenne, mais qui peut atteindre 3,3 ha (Hallworth et al., 2008b). L'étendue du domaine vital des mâles territoriaux en Alberta varie entre 0,18 et 9,87 ha (moyenne = 1,43 ha) (Lesser Slave Lake Bird Observatory et Université de l'Alberta, données inédites). Les habitats locaux de l'espèce présentent généralement des arbustes et une couverture au sol abondants, des arbres où les mâles peuvent se percher pour chanter, et un couvert forestier bas (Hobson et Bayne, 2000a; Hobson et Bayne, 2000b; Hallworth et al., 2008b, Chace et al., 2009). La présence d'un tapis forestier complexe (comprenant par exemple des arbres couchés et de petites buttes), de fougères et d'arbustes offre probablement un couvert protecteur pour les nids et les jeunes (Chace et al., 2009).
Des Parulines du Canada sont souvent observées près d'eaux stagnantes ou de cours d'eau (Reitsma et al., 2010; Ball et al., 2013). Dans une étude menée dans le nord du Minnesota, il a été observé que les Parulines du Canada étaient le plus communes à moins de 100 m des corridors riverains (Zlonis et Niemi, 2014), et Ball et al.(2013) ont observé qu'à l'échelle locale, les vieilles forêts de feuillus sises à proximité de petits cours d'eau encaissés offraient un habitat convenable à l'espèce en Alberta.
Sites de nids
Les nids sont construits sur le sol ou à proximité (Reitsma et al., 2010). Ils sont aménagés sur de la mousse et des buttes, ainsi que dans des trous parmi des masses de racines, des souches d'arbres en décomposition, des touffes de graminées, des cavités rocheuses, etc. (Reitsma et al., 2010). Ils sont aussi généralement aménagés (et réussissent significativement mieux) dans des zones renfermant des débris ligneux grossiers et une forte densité de tiges de plantes ligneuses et de fougères, et où ils peuvent être bien dissimulés (Hallworth et al., 2008b; Chace et al., 2009; Goodnow et Reitsma, 2011).
Alimentation
Les adultes s'alimentent de divers insectes et araignées principalement dans la strate arbustive, surtout en capturant au vol ou en glanant leurs proies, mais aussi ailleurs (Sodhi et Paszkowski, 1995). Parmi leurs proies, on compte des moustiques, des mouches, des lépidoptères, des coléoptères, des vers, des abeilles et des gastéropodes (Reitsma et al., 2010).
Les adultes nourrissent leurs oisillons et leurs jeunes d'aliments divers, comme des chenilles, des papillons, des moustiques et des tipules (Bent, 1953; Reitsma et al., 2010). Les jeunes restent près du nid durant plusieurs jours après l'avoir quitté, dans le couvert du sous-étage (Reitsma et al., 2010).
Habitat de migration et d'hivernage
La Paruline du Canada migre durant la nuit. Durant le jour, elle fait halte dans divers milieux qui sont habituellement mouillés et pourvus d'arbustes, notamment dans des forêts, des marécages arborés et des zones riveraines. Reitsma et al. (2010) ont aussi signalé que des Parulines du Canada sont observées durant la migration dans des buissons et des arbustes près des limites de parcs et de zones habitées.
La Paruline du Canada hiverne généralement en forêt pluviale ou en forêt nuageuse à des altitudes de 500 à 2 500 m (COSEPAC, 2008; Bakermans et al., 2009; Bakermans et al., 2012; BirdLife International, 2014). En Colombie, l'espèce a été signalée dans des forêts tropicales sèches, des forêts montagnardes, des forêts subtropicales ou submontagnardes humides et des forêts nuageuses, ainsi qu'à des sites perturbés, comme des plantations forestières d'essences mélangées, des systèmes agroforestiers et des plantations sous ombrage (BirdLife International, 2014). En Équateur, l'espèce a été le plus souvent signalée dans des forêts humides matures et de seconde venue (BirdLife International, 2014).
Durant l'hiver, il ne semble pas que l'espèce soit territoriale; en effet, on la voit souvent s'alimenter en groupes lâches comprenant plusieurs autres espèces de passereaux (COSEPAC, 2008; Reitsma et al., 2010).
Facteurs limitatifs
Les facteurs limitatifs influent sur la survie et la reproduction d'une espèce, et donc sur ses effectifs. Par exemple, le séjour des Parulines du Canada dans leurs lieux de reproduction est de courte durée comparativement à celui des autres parulines, ce qui impose des contraintes particulièrement fortes pour ce qui est de la durée et du moment de la reproduction, ainsi que du moment de la migration (Flockhart, 2010; Reitsma et al., 2010). Ce facteur réduit la possibilité pour l'espèce de produire plus d'une couvée par saison et de s'adapter aux changements climatiques, ce qui limite sa capacité de réagir aux menaces et d'éventuellement se rétablir une fois celles ci atténuées ou éliminées.
4. Menaces
4.1 Évaluation des menaces
| Catégorie de la menace | Menace | Niveau de préoccupationNote de bas de paged | Étendue | Occurrence | Fréquence | GravitéNote de bas de pagee | Certitude causaleNote de bas de pagef |
|---|---|---|---|---|---|---|---|
| Perte ou dégradation de l'habitat | Habitat autre que l'habitat de reproduction : déforestation et conversion des terres | Élevé | Généralisée | Courante | Continue | Élevée | Moyenne |
| Perte ou dégradation de l'habitat | Habitat de reproduction : enlèvement d'arbustes | Élevé | Généralisée | Courante | Continue | Modérée | Moyenne |
| Perte ou dégradation de l'habitat | Habitat de reproduction : conversion des terres | Élevé | Généralisée | Courante | Continue | Modérée | Moyenne |
| Perte ou dégradation de l'habitat | Habitat de reproduction : exploitation forestière | Moyen | Généralisée | Courante | Continue | Modérée | Moyenne |
| Perte ou dégradation de l'habitat | Exploration et exploitation de ressources énergétiques et minières | Moyen | Généralisée | Courante | Continue | Modérée | Faible |
| Perte ou dégradation de l'habitat | Broutage excessif | Moyen | Localisée | Courante | Continue | Modérée | Moyenne |
| Modifications de la dynamique écologique ou des processus naturels | Disponibilité réduite d'insectes proies (causes ultimes : perte de milieux producteurs d'insectes, désynchronisation des périodes de reproduction de l'espèce et d'abondance de ses proies à cause des changements climatiques, acidification des milieux et pesticides) | Moyen | Généralisée | Courante | Continue | Modérée | Moyenne |
| Mortalité accidentelle | Collisions contre des structures anthropiques et des véhicules | Élevé | Généralisée | Courante | Saisonnière | Modérée | Moyenne |
| Espèces ou génomes exotiques, envahissants ou introduits | Espèces indigènes ou non indigènes problématiques | Faible | Localisée | Courante | Continue | Faible | Faible |
| Climat et catastrophes naturelles | Déplacement et altération de l'habitat | Inconnu | Généralisée | Courante | Continue | Inconnue | Faible |
| Climat et catastrophes naturelles | Températures extrêmes et tempêtes | Inconnu | Généralisée | Courante | Saisonnière | Inconnue | Faible |
| Pollution | Pesticides (effets directs) | Inconnu | Localisée | Courante | Saisonnière | Inconnue | Faible |
| Pollution | Mercure | Inconnu | Généralisée (dans l'est du Canada) |
Courante | Continue | Inconnue | Faible |
| Pollution | Précipitations acides | Inconnu | Généralisée (dans l'est du Canada) |
Courante | Continue | Inconnue | Faible |
4.2 Description des menaces
Les menaces sont décrites ici dans l'ordre où elles sont présentées dans le tableau 2. Selon la Boreal Songbird Initiative (initiative des oiseaux chanteurs de la zone boréale), 24 % de l'habitat de reproduction de la Paruline du Canada se trouvant dans la forêt boréale du Canada a été perturbé par des activités humaines (Boreal Songbird Initiative, 2012), mais la mesure dans laquelle ces perturbations affectent directement les effectifs, la survie et la productivité de l'espèce n'a pas été quantifiée.
La majeure partie de l'information concernant les menaces pesant sur la Paruline du Canada est issue d'études qui ont été menées dans ses lieux de reproduction. Cependant, les menaces existant dans les endroits que l'espèce fréquente en dehors de la période de reproduction peuvent être particulièrement importantes pour elle (Reitsma et al., 2010).
Habitat autre que l'habitat de reproduction : déforestation et conversion des terres
Il a été avancé que la déforestation intensive dans les quartiers d'hiver de la Paruline du Canada pourrait être une menace importante pour l'espèce (COSEPAC, 2008; BirdLife International, 2014). Cette hypothèse est de façon générale simplement fondée sur les niveaux importants de déforestation dans ces régions, plutôt que sur l'observation d'une relation causale directe entre la population de Parulines du Canada et cette menace (BirdLife International, 2014).
En 1991, il a été établi que la superficie boisée dans le nord des Andes (Pérou, Équateur et Colombie) avait diminué d'environ 90 % par rapport à ses niveaux historiques (Henderson et al., 1991), et il a été estimé qu'en 1998, la superficie des forêts andines de Colombie qui avait été déboisée pour l'agriculture avait atteint 180 600 km² (69 %) (Etter et al., 2006). Bien qu'il puisse y avoir eu localement certains gains au cours de la dernière décennie (Sánchez-Cuervo et al., 2012), de vastes étendues de forêt se trouvant dans l'aire d'hivernage de l'espèce (particulièrement en Colombie) continuent de présenter des tendances globales de superficie boisée à la baisse (Portillo-Quintero et al., 2012; Hansen et al., 2013).
Les causes ultimes de la déforestation, qui varient selon les endroits, comprennent l'empiètement humain, l'accroissement des superficies de pâturage, la conversion des cultures de café sous ombrage en cultures de café tolérant l'ensoleillement, la récolte de bois, les plantations de fruits indigènes (narangille), d'autres activités agricoles, et les monocultures (Davis et al., 1997; Portillo-Quintero et al., 2012; BirdLife International, 2014).
Une étude menée en Colombie a montré que la probabilité de présence de la Paruline du Canada a diminué avec la perte de diversité structurale des milieux (Suarez et Colorado (2013) dans BirdLife International, 2014)). McDermott et Rodewald (2014) ont observé que les Parulines du Canada étaient le plus nombreuses dans les forêts et les plantations de cardamome sous ombrage, moins nombreuses dans les plantations de café sous ombrage, et le moins nombreuses dans les systèmes sylvopastoraux, ce qui correspond à un gradient décroissant de fermeture du couvert forestier, de complexité verticale, de complexité de l'étage intermédiaire, et de complexité de l'étage supérieur.
Il y a aussi eu d'importantes pertes de forêt entre 2000 et 2012 dans de vastes portions de l'aire de migration de l'espèce, particulièrement en Amérique centrale (Hansen et al., 2013). Cependant, il a été observé que les Parulines du Canada fréquentent les strates forestières inférieures à feuillage dense durant la migration au Costa Rica (Wolfe et al., 2014). On trouve souvent ce type de sous-étage complexe dans les forêts secondaires jeunes, ce qui a conduit Wolfe et al. (2014) à avancer que les migrateurs pourraient ne pas manquer de bonnes haltes migratoires en Amérique centrale, où beaucoup de forêts primaires sont converties en forêts secondaires.
Habitat de reproduction : enlèvement d'arbustes
La présence d'un couvert arbustif dense est considérée comme une caractéristique essentielle de l'habitat de reproduction de la Paruline du Canada (voir la section 3.3, Besoins de la Paruline du Canada). Par conséquent, les activités qui enlèvent ou détruisent le couvert arbustif peuvent rendre le milieu inhospitalier. Par exemple, les arbustes peuvent être détruits (à dessein ou indirectement) par certaines pratiques sylvicoles (p. ex. application d'herbicides, désherbage, éclaircie) ou durant la récolte (p. ex. récolte sélective, coupe à blanc) (Cooper et al., 1997; Schieck et al., 2000). Les dommages peuvent être réduits grâce à des pratiques qui préservent un sous-étage complexe et une couverture au sol dense, dont a besoin l'espèce, mais les coupes à blanc et la régénération en révolution courte de peuplements équiennes ne reproduisent pas les processus naturels (p. ex. incendie, infestations d'insectes, chablis) qui préserveraient normalement les caractéristiques d'habitat existant dans les forêts boréales canadiennes naturelles (Bergeron et Fenton, 2012). Les efforts visant à réduire les effets néfastes sur le sous-étage arbustif seraient bénéfiques pour la Paruline du Canada (dans la mesure où les autres caractéristiques de son habitat sont aussi conservées), mais la présence d'une strate arbustive dense et abondante peut aussi ralentir la succession forestière dans certaines forêts (p. ex. les forêts d'épinettes noires (Thiffault et al., 2013)).
Habitat de reproduction : conversion des terres
La conversion des terres a été intensive dans certaines parties de l'aire de reproduction de la Paruline du Canada, particulièrement dans la zone de la forêt feuillue nordique et la zone de la forêt mixte boréale. Par exemple, 73 % de la forêt mixte boréale en Saskatchewan a été transformée pour faire place à l'agriculture, 25 % de cette forêt ayant été perdue entre 1966 et 1994 (Hobson et al., 2002). Young et al. (2006) ont calculé un taux annuel de changement de la couverture forestière le long de la bordure sud de la zone boréale albertaine de -0,82 %; or, selon Ball et al. (2013), la Paruline du Canada serait abondante par endroits dans cette région.
Les baisses d'effectifs de la Paruline du Canada dans le nord est des États-Unis de 1950 à 1980 pourraient avoir été causées par le drainage de forêts marécageuses pour l'agriculture et le développement urbain (Miller, 1999), mais les preuves à cet effet sont indirectes. Hobson et Bayne (2000c) ont observé que l'espèce était plus commune en forêt continue que dans des fragments de forêt entourés de terres agricoles, et se tenait bien à l'intérieur des forêts et non à leurs lisières. En Alberta, il a été observé que la présence de terres cultivées et de zones urbaines ou industrielles s'accompagnait d'une réduction des densités de Parulines du Canada à l'échelle locale, mais non à l'échelle du paysage (Ball et al., 2013). Les impacts directs de la conversion des terres sur les effectifs de l'espèce sont inconnus.
Habitat de reproduction : exploitation forestière
Au Canada, les taux de récolte sont les plus élevés au Québec, en Colombie-Britannique et en Ontario. Les taux de récolte semblaient relativement stables au Canada entre les années 1980 et 2008 (Maseket al., 2011), mais ont diminué depuis 2008 (BDNF, 2014). De 2000 à 2012, environ 11 041 217 ha de forêt ont été récoltés dans l'ensemble du Canada (BDNF, 2014). Une bonne partie de l'aire de reproduction de la Paruline du Canada fait l'objet d'activités d'aménagement forestier, mais les pratiques forestières varient dans l'ensemble de cette aire.
De façon générale, la récolte forestière peut avoir des impacts négatifs à court terme sur les oiseaux durant leur nidification en perturbant les activités de reproduction (Hobson et al., 2013). Les nids ou les œufs peuvent être détruits ou perturbés par mégarde durant la coupe d'arbres et d'autres végétaux (p. ex. éclaircie précommerciale) (Environment Canada, 2014). Les activités perturbant les nicheurs peuvent aussi entraîner l'échec de leur nidification (Environment Canada, 2014). Hobson et al. (2013) ont estimé qu'entre 616 000 et 2,09 millions de nids (de nombreuses espèces) sont perdus chaque année à cause de la récolte forestière industrielle.
Des études menées dans certaines parties de l'aire de reproduction de la Paruline du Canada (particulièrement dans l'est) ont révélé que l'espèce pourrait tolérer un certain niveau de récolte forestière (Hagen et al., 1997; King et DeGraaf, 2000; Chace et al., 2009). Les régimes de récolte dont les impacts reproduisent à peu près ceux de phénomènes comme les tempêtes, les incendies et les infestations d'insectes, qui modifient de façon naturelle le milieu, peuvent fournir un habitat convenable pour la reproduction de la Paruline du Canada. On a observé que l'espèce était relativement nombreuse à certains sites de 5 à 20 ans après récolte (coupes partielles, coupes progressives et coupes à blanc), quand certains arbres de l'étage supérieur étaient conservés et le sous-étage était relativement dense (Titterington et al., 1979; Hagen et al., 1997; King et DeGraaf, 2000; DeGraaf et Yamasaki, 2003; Chace et al., 2009). Hallsworth et al. (2008b) n'ont pas observé de différence significative entre les succès reproducteurs de mâles en parcelles de seconde venue et en marécages à érable rouge au New Hampshire. Ces auteurs ont avancé que l'habitat de la Paruline du Canada pourrait être maintenu à l'échelle du paysage grâce à une rotation des récoltes veillant à laisser des arbres sur pied et à créer des sous-étages denses (Hallworth et al., 2008b). Cependant, cette façon de faire pourrait n'être appropriée que dans certaines parties de l'aire de reproduction de l'espèce. Dans le sud de la forêt boréale du Québec, la Paruline du Canada est surtout observée dans des zones d'arbustes feuillus denses le long des emprises créées par l'industrie du bois (B. Drolet, comm. pers.)
Dans certaines parties de son aire de reproduction, la Paruline du Canada préfère les vieilles forêts (âgées de plus de 120 ans, et souvent perturbées par des processus naturels) (Drapeau et al., 2000; Schieck et Song, 2006; Zlonis et Niemi, 2014). Par exemple, une étude menée dans le nord du Minnesota a révélé que l'espèce était plus commune en forêt sauvage qu'en forêt aménagée (Zlonis et Niemi, 2014). Haché et al. (2014) ont observé que les densités de Parulines du Canada étaient généralement plus élevées dans les zones renfermant des arbres de grande taille (caractéristique servant d'indicateur de l'âge de la forêt), et DeGraaf et al. (1991) que l'espèce était plus nombreuse dans des peuplements non éclaircis que dans des peuplements éclaircis. On a constamment trouvé de plus fortes densités dans les blocs d'inventaire ayant un moindre pourcentage de superficies récoltées en Alberta, mais les résultats n'étaient pas toujours significatifs (Lesser Slave Lake Bird Observatory et Université de l'Alberta, données inédites). Les récoltes à courte rotation qui ne permettent pas l'établissement de forêts âgées structuralement complexes pourraient avoir un impact négatif sur la Paruline du Canada dans les régions où celle-ci préfère les vieilles forêts. Selon Angers et al. (2005), il est très peu probable que les coupes de jardinage permettraient le développement de caractéristiques propres aux vieilles forêts dans les intervalles actuellement observés entre les coupes, et ces coupes pourraient avoir des impacts négatifs sur la disponibilité de débris ligneux et les caractéristiques de ces débris (facteurs importants pour le succès de reproduction des Parulines du Canada).
Ball et al. (2013) ont observé des Parulines du Canada dans des milieux de début de succession en Alberta, mais ces individus se trouvaient dans des parcelles de forêt résiduelle (et non pas dans des zones renfermant seulement des arbres résiduels – comme on l'avait rapporté ailleurs (p. ex. Hallworth et al., 2008b)). Cette observation a conduit Ball et al. (2013) à avancer qu'il faudrait maintenir de vastes peuplements de vieille forêt feuillue (particulièrement au voisinage de cours d'eau), et accroître la largeur des zones tampons riveraines dans lesquelles la forêt doit être conservée.
Exploration et exploitation de ressources énergétiques et minières
La recherche de ressources énergétiques (p. ex. pétrole, gaz et ressources hydrauliques) et de minéraux, l'exploitation de ces ressources (p. ex. inondation de vastes étendues pour créer des réservoirs et résidus miniers), et l'aménagement de corridors de transport (p. ex. pipelines, lignes électriques et routes) ont causé une perte, une dégradation et une fragmentation importantes d'habitat dans certaines parties de l'aire de répartition de la Paruline du Canada (Drummond et Loveland, 2010; Masek et al., 2011; Birch et Kaye, 2012). Les activités associées à ces industries peuvent aussi entraîner une destruction non intentionnelle de nids, d'œufs, d'oisillons ou d'adultes (Van Wilgenburg et al., 2013).
Van Wilgenburg et al.(2013) ont estimé qu'environ 48 400 ha sont perturbés chaque année par l'aménagement de puits, de pipelines et de lignes sismiques au sein de l'écozone boréale dans le bassin sédimentaire de l'Ouest canadien. Il en découle la perte d'environ 7 301 nids (de nombreuses espèces) dans cette région chaque année (Van Wilgenburg et al., 2013). Van Wilgenburg et al. (2013) ont estimé que ces aménagements faisaient perdre assez peu de recrues (52-107) à la population de Parulines du Canada, en raison des densités relativement faibles de l'espèce. L'aménagement de puits, de pipelines et de lignes sismiques est particulièrement intense dans le nord de l'Alberta et le nord-est de la Colombie-Britannique (Schneider et al., 2003; Calvert et al., 2013; Van Wilgenburg et al., 2013). Des activités pétrolières et minières et l'aménagement connexe de pipelines et de routes ont aussi cours localement dans les quartiers d'hiver de l'espèce du nord des Andes (Davis et al., 1997; BirdLife International, 2014).
Des activités minières ciblant divers minéraux (p. ex. or, diamant, zinc, plomb et cuivre) ont cours un peu partout dans l'aire de répartition canadienne de la Paruline du Canada (Stothart, 2011).La superficie totale visée par des baux d'exploitation minière au Canada s'élève à 2,1 millions d'hectares (Cheng et Lee, 2014), ce qui équivaut à environ 0,21 % de la superficie totale du Canada. sont l'Alberta (3 206 km²), l'Ontario (1 686 km²) et le Manitoba (1 463 km²) (Cheng et Lee, 2014). L'impact de ces industries sur la population de Parulines du Canada est inconnu.
L'exploitation de ressources énergétiques et minières peut altérer les régimes hydrologiques locaux et le niveau d'humidité des sols, qu'on pense au prélèvement direct d'eau (p. ex. pour la fracturation) et aux installations hydroélectriques (p. ex. barrages). Comme les Parulines du Canada sont plus nombreuses près des corridors riverains dans l'ensemble de l'aire de l'espèce (Reitsma et al., 2010; Zlonis et Niemi, 2014) et se reproduisent notamment dans des marécages arborés, des tourbières et près d'eaux libres dans l'est (Reitsma et al. 2010), elles seraient probablement affectées par des changements des régimes hydrologiques.
Broutage excessif
Le broutage exercé par les cerfs peut altérer radicalement la strate arbustive utilisée par les Parulines du Canada pour nicher et s'alimenter (Reitsma et al., 2010) en réduisant la quantité et la diversité d'arbustes et en modifiant la dynamique de la végétation (Côté et al., 2004; Rooney, 2009). Une étude menée dans le nord est des États-Unis a montré que les effectifs de Parulines du Canada étaient inférieurs dans les zones renfermant de forts effectifs de cerfs de Virginie (Odocoileus virginianus) (DeGraaf et al., 1991). Comme la Paruline du Canada, la Paruline du Kentucky (Geothlypis formosus) se reproduit principalement dans le sous-étage (McDonald, 2013). McShea et al. (1995) ont montré que les Parulines du Kentucky étaient observées plus fréquemment dans les zones où les effectifs de cerfs de Virginie étaient faibles. Le broutage exercé par les cerfs a aussi affecté des espèces européennes ayant des besoins en matière d'habitat semblables à ceux de la Paruline du Canada. Par exemple, Holt et al. (2013) ont démontré que des Fauvettes à tête noire (Sylvia atricapilla) établissaient leur territoire plus tôt et présentaient un meilleur état corporel en l'absence de cerfs. Cela s'explique par le fait que les cerfs broutent la strate arbustive dont les oiseaux ont besoin (Reitsma et al., 2010). Les populations de cerfs de Virginie sont en augmentation dans la partie sud de l'aire de la Paruline du Canada (Russell et al., 2001). Pour le moment, on ne connaît pas très bien l'étendue de cette menace ni le niveau de préoccupation qui s'y rattache pour la population de Parulines du Canada, mais le problème devrait être le plus marqué dans le nord est des États-Unis et le sud est du Canada, où les cerfs de Virginie sont particulièrement abondants (Russell et al., 2001).
Le broutage non contrôlé exercé par le bétail a aussi contribué à la dégradation de l'habitat forestier de la Paruline du Canada dans les aires de migration, d'hivernage et de reproduction de l'espèce. Du bétail broute le long de la bordure méridionale de la zone boréale dans l'Ouest canadien (principalement dans les tremblaies), dans des secteurs qui conviendraient autrement à la Paruline du Canada. Les effets du broutage exercé par le bétail sur l'habitat dépendent du moment où il est effectué et de sa durée, de même que des propriétés du sol (Krzic et al., 2004).
Réduction de la disponibilité d'insectes proies
Les populations d'insectivores aériens connaissent des baisses marquées, particulièrement dans le nord est de l'Amérique du Nord (Nebel et al., 2010). Toutes les espèces de ce groupe varié d'oiseaux ont pour point commun de consommer des insectes, ce qui a conduit de nombreux chercheurs à penser qu'une réduction de la disponibilité d'insectes proies dans les aires de reproduction, de migration ou d'hivernage pourrait probablement contribuer aux tendances à la baisse des populations de ces oiseaux (Nebel et al., 2010; Paquette et al., 2014). Bien que la Paruline du Canada ne soit pas considérée comme étant un insectivore aérien, elle s'alimente principalement d'insectes (souvent en vol); par conséquent, les effets des changements de disponibilité d'insectes sur la Paruline du Canada pourraient être semblables à ceux soupçonnés pour les insectivores aériens.
Les populations d'insectes connaissent des baisses significatives partout dans le monde. Un examen récent des tendances mondiales des populations fauniques a révélé que 33 % de tous les insectes dont l'UICN a évalué les tendances connaissent une baisse d'effectif, et bon nombre connaissent aussi un rétrécissement de leur aire de répartition (Dirzo et al., 2014). Ce phénomène est observé à l'échelle mondiale, mais les baisses sont plus prononcées dans les régions fortement perturbées, notamment sous les tropiques (Dirzo et al., 2014). Les causes possibles de la réduction de la disponibilité d'insectes proies sont décrites ci dessous.
Perte de milieux producteurs d'insectes
Nombre d'insectes sont confinés à des milieux particuliers durant au moins une partie de leur cycle biologique, de sorte que toute activité qui affecte ces milieux peut nuire à ces insectes. Plus de 90 % des groupes d'insectes considérés comme menacés sont touchés par la perte ou la dégradation de leur habitat (Price et al., 2011). Diverses activités humaines altèrent ou détruisent les milieux naturels dont ont besoin certains stades de développement des insectes, comme le drainage de milieux humides et l'extraction de tourbe, l'agriculture intensive, la destruction de milieux humides, des activités industrielles et le développement urbain (U.S. Bureau of Land Management, 1978; Benton et al., 2002; Price et al., 2011; Brooks et al., 2012). Par exemple, les activités de forage, de construction et d'aménagement associées à l'exploitation gazière et pétrolière peuvent mener à la perte de l'habitat des insectes et donc entraîner une réduction des populations et altérer la composition des assemblages d'insectes (U.S. Bureau of Land Management, 1978). Selon Foster (1991), le drainage des milieux humides et l'extraction de tourbe constituent une menace importante pour les populations d'insectes.
Les effets de la perte de l'habitat des insectes sur la Paruline du Canada ne seraient pas limités à son aire de reproduction; ils pourraient aussi se faire sentir dans l'aire autre que l'aire de reproduction. La fragmentation des forêts pluviales pourrait entraîner une baisse de la richesse en espèces d'insectes et des populations de certaines espèces (Price et al., 2011). En général, les réponses des insectes aux changements d'utilisation des terres dans les Andes tropicales sont tributaires du contexte, mais certaines recherches ont montré que la richesse spécifique, la diversité d'espèces et les effectifs diminuent par suite de perturbations associées à l'utilisation des terres (Larsen et al., 2011).
Désynchronisation des périodes de reproduction de la Paruline du Canada et d'abondance des proies
Chez les oiseaux, il y a souvent synchronisation entre la période de reproduction (plus précisément l'éclosion des œufs) et la période où la nourriture disponible est le plus abondante, mais les changements climatiques ont fait que les pics d'abondance de certains insectes ont été devancés (Both et al., 2009). Comme le réchauffement climatique est moins marqué dans les quartiers d'hiver des Parulines du Canada que dans leurs lieux de reproduction, les signaux les incitant à entreprendre leur migration vers le nord pourraient se manifester trop tard pour qu'elles puissent arriver à leurs lieux de reproduction au moment optimal (Jones et Cresswell, 2010). Par conséquent, les changements climatiques font qu'il y a désynchronisation entre la reproduction et l'abondance maximale de proies (insectes) pour les espèces qui ne s'adaptent pas à l'évolution du climat au même rythme que leurs proies (Strode, 2003). Both et al.(2006) ont observé qu'aux Pays Bas, un insectivore aérien, le Gobemouche noir (Ficedula hypoleuca), a connu une baisse d'effectif de 90 % de 1987 à 2003 dans des régions où le pic d'abondance de ses proies survenait trop tôt dans la saison de reproduction pour que les parents puissent nourrir adéquatement leurs oisillons. Strode (2003) a observé que de nombreuses parulines qui se nourrissent de chenilles pourraient ne pas s'être adaptées aux changements climatiques en migrant plus tôt. Par conséquent, elles ne trouveraient pas suffisamment de nourriture pour se rétablir de leur migration, se reproduire et nourrir leurs oisillons. On a aussi observé chez des Mésanges charbonnières (Parus major) une éclosion des œufs ne se produisant pas au moment du pic de biomasse de chenilles du fait du réchauffement climatique récent (Visser et al., 2006). Ce décalage a influé sur le poids des oisillons et le nombre d'entre eux ayant atteint l'âge de l'envol (Visser et al., 2006). La désynchronisation entre la reproduction des oiseaux et la disponibilité des proies a aussi été associée aux baisses d'effectifs d'oiseaux migrateurs d'Europe (Møller et al., 2008; Saino et al., 2011), et contribuerait aux baisses d'effectifs d'autres espèces aviaires fortement tributaires d'invertébrés, comme le Quiscale rouilleux (Euphagus carolinus) (McClure et al., 2012).
Les populations d'oiseaux migrateurs effectuant de longues migrations et se reproduisant dans des habitats saisonniers sont plus vulnérables aux changements climatiques parce que la désynchronisation temporelle est plus probable et plus marquée (Both et al., 2006; Both et al., 2009). Comme la Paruline du Canada est un insectivore, migre sur de longues distances et se reproduit et s'alimente dans des habitats saisonniers, il est certainement plausible que les changements climatiques induisent une désynchronisation entre sa reproduction et la disponibilité de proies, mais on ne dispose pas actuellement de données propres à l'espèce à ce sujet.
Acidification de l'habitat
Depuis les années 1980, le taux de dépôt acide a grandement diminué, mais des composés acidifiants (p. ex. dioxyde de soufre et oxydes d'azote) sont encore libérés dans l'environnement (Shannon, 1999; Environnement Canada, 2014b). L'acidification des eaux de surface peut réduire les effectifs et la diversité des insectes volants dont le cycle biologique comporte une phase aquatique (Graveland, 1998), et l'acidification des forêts peut éclaircir le couvert forestier et entraîner ainsi des changements des communautés et des effectifs d'insectes (Mahony et al., 1997; Brotons et al., 1998; Zang, 1998). Certaines proies de la Paruline du Canada, comme les moustiques (Culicidés), les tipules (Tipulidés), les chenilles (Lépidoptères) et les araignées (Arachnides), peuvent être affectées par l'acidification de l'habitat. Il a été observé que la réduction du succès reproducteur d'Hirondelles bicolores (Tachycineta bicolor) vivant près de milieux humides acidifiés en Ontario était liée à des changements de la disponibilité de proies riches en calcium pour les oisillons (Blancher et McNicol, 1991), et que l'acidification des forêts était impliquée dans la baisse d'effectif de la Grive des bois (Hylocichla mustelina) (Hames et al., 2002). Cependant, une étude menée dans le centre de l'Ontario a montré que la productivité de passereaux occupant des sites acidifiés n'était pas différente de celle de passereaux occupant des sites non acidifiés (Mahony et al., 1997). Pour le moment, rien n'indique que l'acidification de l'habitat réduirait les effectifs ou la qualité nutritive des insectes proies dans l'ensemble de l'aire de la Paruline du Canada, mais ce phénomène pourrait toucher l'espèce dans les endroits recevant d'importants dépôts acides, ainsi que dans l'est de l'Amérique du Nord, où le pouvoir tampon des sols est relativement faible.
Pesticides (effets indirects)
La plupart des pesticides organochlorés (dont le dichlorodiphényltricholoroéthane – DDT) sont interdits en Amérique du Nord depuis des décennies, mais des données indiquent que des migrateurs insectivores néotropicaux sont encore exposés à ce type de pesticides dans l'ensemble de leurs aires de répartition (Sager, 1997; Klemens et al., 2000). Par ailleurs, ces substances chimiques peuvent avoir des effets à long terme sur les communautés d'insectes, et donc sur les oiseaux insectivores. Des données historiques ont révélé une baisse marquée des coléoptères et un accroissement des punaises (Hémiptères) dans le régime alimentaire du Martinet ramoneur (Chaetura pelagica), qui étaient temporellement corrélés avec une forte hausse de la présence de DDT et de ses métabolites dans l'environnement. Nocera et al. (2012) ont avancé que le DDT avait entraîné des baisses des populations de coléoptères et des modifications très marquées (et possiblement permanentes) des communautés d'insectes, ce qui aurait appauvri en nutriments le régime alimentaire des Martinets ramoneurs et ainsi contribué à leurs baisses d'effectifs.
Les effets néfastes des insecticides chimiques ont suscité l’utilisation accrue d’insecticides biologiques. Actuellement, les insecticides utilisés dans les activités forestières au Canada sont principalement biologiques (Bacillus thuringiensis var. kurstaki (Btk)) et ciblent des larves de lépidoptères, comme la tordeuse du pin gris (Choristoneura pinus) et la tordeuse des bourgeons de l’épinette (Choristoneura fumiferana). La superficie annuelle moyenne traitée avec le Btk dans les forêts canadiennes de 1988 à 2000 a été de 273 440 ha (fourchette : 73 209 – 855 535 ha) (BDNF, 2014). En 2012, du Btk a été appliqué dans les forêts de quatre provinces canadiennes : le Québec (98 044 ha), le Manitoba (828 ha), la Saskatchewan (15 639 ha), et la Colombie-Britannique (116 012 ha) (BDNF, 2014). En moyenne, c’est le Québec qui traite la plus importante superficie de forêt avec le Btk par année (1988-2012) (BDNF, 2014). Bon nombre d’insecticides microbiens sont considérés comme non toxiques pour les oiseaux, et les données concernant leurs possibles effets indirects par l’entremise de changements de la disponibilité de proies demeurent non concluantes. Dans un secteur de 12 803 ha de l’île de Vancouver, en Colombie-Britannique, on n’a observé aucune différence de richesse spécifique ou d’abondance relative chez les passereaux un an après pulvérisation de Btk contre la spongieuse (Lymantria dispar) (Sopuck et al., 2002). Holmes (1998) a observé que le traitement au Btk de sites ontariens n’a pas affecté la survie et la croissance d’oisillons de Parulines obscures (Vermivora peregrina); bien que les nids se trouvant dans les sites traités ont présenté des couvées et des nichées de plus petite taille et des taux d’éclosion inférieurs, les différences n’étaient pas significatives. D’autres études ont révélé que des pesticides microbiens avaient sur des oiseaux des impacts indirects significatifs. Dans un secteur traité au Btk en Ontario, des oisillons de Tétras du Canada (Dendragapus canadensis) ont présenté des taux de croissance significativement inférieurs à ceux d’oisillons élevés dans des sites d’étude non traités au Btk (Norton et al., 2001). Norton et al. (2001) ont attribué ce résultat à la réduction de la disponibilité de larves de lépidoptères par suite de la pulvérisation.
Les insecticides néonicotinoïdes ont été introduits dans les années 1990, et bien que leurs taux d'utilisation soient mal connus dans l'aire de la Paruline du Canada, on estime que près de 11 millions ha de terres cultivées sont traités avec ce type de produits dans les Prairies canadiennes (Main et al., 2014). Les néonicotinoïdes sont généralement utilisées dans les terres agricoles, mais on en a détecté dans des milieux humides (Main et al., 2014) et des cours d'eau au Canada (Environnement Canada, 2011a; Xing et al., 2013). Les impacts indirects des néonicotinoïdes sur les Parulines du Canada sont inconnus, mais comme l'habitat de l'espèce ne comprend pas de terres cultivées, ils sont probablement très faibles, même si ces insecticides sont mobiles et persistants dans l'environnement (Hladik et al., 2014). Néanmoins, Mineau et Palmer (2013) ont laissé entendre que les effets des néonicotinoïdes sur les oiseaux pourraient déborder des terres cultivées et s'étendre à l'échelle des bassins hydrographiques ou à l'échelle régionale; ainsi, comme les néonicotinoïdes peuvent avoir des impacts sur les insectes et les oiseaux vivant en dehors des terres arables, nous les avons inclus parmi les pesticides constituant une menace pour la Paruline du Canada du fait d'effets indirects. Les néonicotinoïdes ont un impact négatif sur les populations d'insectes, au point qu'en 2013, l'Autorité européenne de sécurité des aliments a déclaré qu'ils présentent un risque « inacceptable » pour les insectes (Goulson, 2014). Aux Pays Bas, les concentrations de néonicotinoïdes dans les eaux de surface ont été corrélées avec les baisses des effectifs d'oiseaux insectivores des terres agricoles (Hallmann et al., 2014). Hallmann et al. (2014) ont laissé entendre que ces baisses découlaient probablement d'une réduction des quantités d'insectes proies par suite de l'utilisation d'insecticide. Les effets indirects de ces insecticides ont aussi été signalés chez l'Alouette des champs (Alauda arvensis), le Bruant jaune (Emberiza citrinella), le Tarier des prés (Saxicola rubetra), le Bruant des roseaux (Emberiza schoeniclus) et le Bruant proyer (Miliaria calandra) (Boatman et al., 2004; Gibbons et al., 2014).
On sait que de grandes quantités de pesticides s'emploient dans les plantations de caféiers situées dans les quartiers d'hiver de la Paruline du Canada (Rappole et al., 2003). Leurs effets sur la disponibilité des proies sont inconnus.
Collisions contre des structures anthropiques et des véhicules
Durant leur migration nocturne, les Parulines du Canada peuvent percuter des structures artificielles, comme des édifices, des tours de communication, des lignes électriques et des éoliennes. Même si ces collisions sont épisodiques, les mortalités qui en découlent peuvent être importantes à l'échelle de la population.
Chaque année, environ 25 millions d'oiseaux (de nombreuses espèces) meurent au Canada par suite de collisions contre des fenêtres (Machtans et al., 2013), et le nombre s'élève à entre 365 et 988 millions aux États Unis (Loss et al., 2014a). Les Parulines du Canada sont considérées comme très susceptibles de percuter des édifices de grande taille ou de petite taille (Loss et al., 2014a). Pour elles, le risque de collision par rapport à la moyenne des espèces est 17,9 fois plus élevé pour tous les types d'édifices, 46,7 fois plus élevé pour les édifices de petite taille, et 25,8 fois plus élevé pour les édifices de grande taille (Loss et al., 2014a).
Quelque 6,8 millions d'oiseaux (de nombreuses espèces) sont tués chaque année par suite de collisions contre des tours de communication aux États-Unis et au Canada (Longcore et al., 2012). Les mortalités sont particulièrement fréquentes pour les migrateurs néotropicaux et les migrateurs nocturnes attirés par les lumières des tours (Longcore et al., 2013). Dans une étude sur les collisions contre les tours de communication dans le centre et l'est de l'Amérique du Nord, on a estimé que 20 622 Parulines du Canada sont tuées chaque année de cette façon (Longcore et al., 2013).
Il a été estimé que, chaque année, de 2,5 à 25,6 millions d'oiseaux (de nombreuses espèces) sont tués par les lignes électriques au Canada (Rioux et al., 2013) et entre 12 et 64 millions aux États-Unis (de 8 à 57 millions par collision, et de 0,9 à 11,6 millions par électrocution) (Loss et al., 2014c). L'ampleur du phénomène pour la Paruline du Canada n'a pas été quantifiée.
Environ 23 300 oiseaux (de nombreuses espèces) sont tués chaque année par suite de collisions contre des éoliennes (Zimmerling et al., 2013). Il a été estimé que près de 50 % de ces mortalités surviendraient en Ontario (Zimmerling et al., 2013). Au Canada, environ 70 Parulines du Canada seraient tuées chaque année par suite de collisions contre des éoliennes (Zimmerling et al., 2013).
Bishop et Brogan (2013) ont estimé qu'environ 3 462 oiseaux (de nombreuses espèces) sont tués par 100 km de route pavée à une ou deux voies à l'extérieur des grands centres urbains au Canada durant chaque saison de reproduction, et Loss et al. (2014b) ont estimé qu'entre 89 et 340 millions d'oiseaux sont tués chaque année aux États-Unis par suite de collisions avec des véhicules. Bien qu'il existe des exceptions, en général, les taux de mortalité par collision avec des véhicules augmentent souvent avec la vitesse du trafic, la largeur des corridors routiers et le niveau des routes par rapport au terrain environnant (Case, 1978; Baudvin, 1997; Loss et al., 2014b). Les passériformes comptent pour 40 % de toutes les mortalités d'oiseaux par collision avec des véhicules en Amérique du Nord, mais la Paruline du Canada ne figure dans aucune des 28 études sur le sujet dont traitent Bishop et Brogan (2013).
Espèces indigènes ou non indigènes problématiques
Le parasitisme des couvées pratiqué par certaines espèces d'oiseaux consiste à pondre des œufs dans le nid d'individus d'une autre espèce, au détriment de ces derniers. Le parasitisme des couvées par les Vachers à tête brune est une menace potentielle pour les Parulines du Canada, au moins par endroits, là où les deux espèces sont présentes (Cooper et al., 1997). Il est établi que des vachers parasitent des nids de Parulines du Canada, mais rien n'indique que cette menace a un impact négatif sur les effectifs de Parulines du Canada (Reitsma et al., 2010).
Les chats domestiques ou harets représentent la plus importante cause de mortalité d'oiseaux liée à l'homme au Canada (Calvert et al., 2013).
On a estimé que dans le sud du Canada, de 2 à 7 % de tous les oiseaux sont tués par des chats annuellement (Blancher, 2013). Ce problème est moins préoccupant dans les régions nordiques et en pleine forêt, mais la Paruline du Canada serait vulnérable à la prédation par les chats (particulièrement parce qu'elle niche près du sol) dans les parties sud et rurales de son aire de reproduction. Elle serait vraisemblablement le plus vulnérable dans son aire de migration, où elle a été signalée près de parcs et de zones habitées.
Le parasitisme des couvées et la prédation peuvent être particulièrement préoccupants pour la Paruline du Canada durant la période de reproduction, du fait qu'elle ne produit qu'une seule couvée.
Déplacement et altération de l'habitat
Les oiseaux migrateurs qui parcourent de longues distances sont tributaires d'habitats multiples dispersés durant leur cycle annuel (habitats de reproduction, de migration et d'hivernage). Cela les rend particulièrement vulnérables aux impacts des changements climatiques, tout changement se produisant dans ces habitats pouvant affecter leurs populations (Newson et al., 2009; Robinson et al., 2009). On dispose de peu d'information qui permettrait d'établir un lien direct entre les changements climatiques et la baisse d'effectif de la Paruline du Canada, mais selon Cumming et al. (2014), les changements climatiques présentent un fort potentiel de modification des répartitions d'oiseaux.
Températures extrêmes et tempêtes
L'augmentation de la fréquence des tempêtes tropicales et d'autres phénomènes météorologiques néfastes (p. ex. fortes pluies, températures extrêmes, tempêtes de vent violentes) causée par les changements climatiques pourrait accroître les échecs de nidification et la mortalité directe sur l'ensemble du cycle annuel de la Paruline du Canada (Rodenhouse et al., 2008). Les épisodes de temps violent pourraient aussi réduire les possibilités d'alimentation, causer des problèmes de thermorégulation pour les adultes et les jeunes, perturber les migrations et endommager les milieux où hiverne l'espèce.
Les incendies sont fortement déterminés par les conditions météorologiques (Flannigan et al., 2009), et l'étendue, l'intensité et la fréquence des incendies de forêt devraient continuer de s'accroître en raison des printemps et étés plus chauds et plus secs (Flannigan et al., 2009; North American Bird Conservation Initiative US Committee, 2010; de Groot et al., 2013; Girardin et al., 2013).
L'impact de ces phénomènes météorologiques extrêmes sur la population de Parulines du Canada est actuellement inconnu.
Pesticides (effets directs)
Selon Mineau et Whiteside (2013), il faudrait porter une attention particulière aux pesticides dans la recherche des facteurs responsables des baisses d'effectifs des populations d'oiseaux en Amérique du Nord, tout spécialement pour les espèces qui se reproduisent ou hivernent dans des zones agricoles, ou qui traversent de telles zones durant leurs migrations. Ces auteurs n'ont pu faire la part entre les effets directs (effets toxiques dus à l'ingestion de produits comme des semences enrobées, à l'inhalation, à l'absorption cutanée, ou à la consommation de proies contaminées) et indirects (par exemple sur l'habitat ou la chaîne alimentaire) des pesticides, et ils ont conclu que les pesticides ont probablement ces deux types d'effets (Mineau et Whiteside, 2013). Il existe très peu de données sur le sujet pour ce qui est de la Paruline du Canada, mais, pour de nombreuses espèces d'oiseaux, des cas de mortalité directe et de perte d'habitat ont été attribués à l'utilisation de pesticides dans les lieux de reproduction et d'hivernage (voir par exemple Chamberlain et al. (2000), Boatman et al. (2004), Mineau (2005)).
La plupart de pesticides organochlorés (dont le DDT) sont interdits depuis des décennies en Amérique du Nord. On ne sait pas très bien dans quelle mesure les Parulines du Canada et d'autres passereaux migrateurs néotropicaux ont été exposés à cette classe de pesticides au fil de leur existence (Gard et al., 1993; Klemens et al., 2000), mais certaines données indiquent que des insectivores migrateurs néotropicaux y sont encore exposés en Amérique du Nord (Sager, 1997; Klemens et al., 2000). Cette exposition peut se produire en toute légalité, du fait d’exceptions dans les dispositions législatives concernant les restrictions d'utilisation de ces pesticides, ou illégalement. Ces pesticides pourraient encore être utilisés en Amérique centrale et en Amérique du Sud (Klemens et al., 2000; Lebbin et al., 2010; Nebel et al., 2010) dans la lutte contre les moustiques nuisibles, ou à des fins agricoles ou autres. L'endosulfan, utilisé principalement dans une grande variété de cultures alimentaires, n'est exceptionnellement pas frappé par l'interdiction concernant les organochlorés, mais son utilisation aux États-Unis fait l'objet d'une élimination progressive qui sera complétée en 2016 parce qu'il a été établi qu'il présente un risque inacceptable pour les travailleurs agricoles et la faune (les oiseaux sont de façon générale passablement vulnérables à l'empoisonnement par l'endosulfan) (U.S. Environmental Protection Agency, 2010). Plusieurs autres pays ont entrepris d'interdire ce pesticide par l'entremise de la Convention de Stockholm sur les polluants organiques persistants, traité international signé en 2001 (Secretariat of the Stockholm Convention, 2011).
Divers composés organophosphorés, dont des organophosphates, et carbamates sont de plus en plus utilisés depuis que l'utilisation de la majorité des pesticides organochlorés a été restreinte en Amérique du Nord dans les années 1970, puis interdite dans les années 1980 (Commission de coopération environnementale de l'Amérique du Nord, 2003). Les oiseaux et autres vertébrés peuvent être affectés s'ils ingèrent ou absorbent de quelque autre façon suffisamment d'organophosphate ou de carbamate, les oiseaux y semblant de façon générale plus sensibles que les autres vertébrés (Freedman, 1995; Friend et Franson, 1999).
Les impacts directs d'une classe relativement nouvelle de pesticides, les néonicotinoïdes, sont inconnus pour les espèces insectivores, comme la Paruline du Canada. Hallmann et al. (2014) ont observé une corrélation entre les concentrations de néonicotinoïdes dans les eaux de surface et des baisses d'effectifs d'oiseaux insectivores aux Pays Bas. Ils ont avancé que ces baisses étaient liées à une réduction de la quantité d'insectes proies, mais ils n'ont pu exclure la possibilité d'effets directs des néonicotinoïdes sur les oiseaux (Hallmann et al., 2014). Le niveau d'exposition de la Paruline du Canada aux pesticides néonicotinoïdes est inconnu, mais, vu les préférences de l'espèce en matière d'habitat, il est probablement faible dans ses lieux de reproduction, même si ces pesticides sont mobiles et persistants dans l'environnement (Hladik et al., 2014).
Dans l'aire d'hivernage de la Paruline du Canada, il se fait dans les plantations de café une forte utilisation de pesticides, lesquels passent dans les eaux de ruissellement (Rappole et al., 2003). On ne sait pas dans quelle mesure cela affecte la Paruline du Canada.
Mercure
Le mercure est un élément présent de façon naturelle dans l'environnement, mais ses concentrations dans celui ci se trouvent accrues par certaines activités humaines. Le transport atmosphérique à longue distance et le dépôt subséquent constituent la principale source de mercure pour de nombreux milieux aquatiques dans une grande partie du paysage (Fitzgerald et al., 1998; U.S. Geological Survey, 2000). Du mercure biodisponible est aussi mobilisé dans les bassins hydrologiques par les activités forestières, la création de réservoirs hydroélectriques et diverses autres activités industrielles (Porvari et al., 2003; Vuori et al., 2003; Wiener et al., 2003). Les concentrations de mercure dans les réseaux trophiques aquatiques sont habituellement corrélées avec de faibles pH, de sorte que dans les réseaux trophiques d'eau douce du Canada, elles vont en s'accroissant d'ouest en est (Depew et al., 2013).
L'exposition au mercure peut réduire le succès reproducteur, altérer la réponse immunitaire, et avoir des effets comportementaux et physiologiques chez les oiseaux (Scheuhammer et al., 2007; Hawley et al., 2009). Des travaux réalisés par Keller et al.(2014) et Rimmer et al. (2010) ont indiqué que le mercure se bioamplifie dans les oiseaux chanteurs terrestres qui consomment des invertébrés. Dans certaines parties de son aire de répartition, la Paruline du Canada pourrait être fortement exposée au méthylmercure (MeHg, forme toxique du mercure) en raison de sa consommation d'insectes prédateurs issus de milieux humides acides, propices à la formation de méthylmercure (Greenberg et Matsuoka, 2010; Evers et al., 2011; Edmonds et al., 2012). Dans une étude récente à grande échelle portant sur le mercure présent dans un oiseau insectivore, le Quiscale rouilleux, on a souligné le danger que peut présenter le mercure pour cet oiseau, particulièrement pour la population du nord est de l'Amérique du Nord (Edmonds et al., 2010). Les concentrations de mercure dans les plumes de Quiscales rouilleux échantillonnées durant la période de reproduction dans l'écorégion de la forêt acadienne, située en Nouvelle-Angleterre et dans les Maritimes (Maine, New Hampshire, Vermont, Nouveau-Brunswick et Nouvelle-Écosse), étaient supérieures de plusieurs ordres de grandeur à celles mesurées dans les plumes échantillonnées dans des quartiers d'hiver du sud des États-Unis et dans des lieux de reproduction de l'Alaska (Edmonds et al., 2010).
Précipitations acides
Les précipitations acides contribuent au déclin des forêts d'épinette et de sapin dans l'est des États-Unis (U.S. Environmental Protection Agency, 2014), et probablement aussi au Canada. L'acidification peut modifier l'habitat en entraînant l'altération des assemblages d'invertébrés du sol (voir la section Réduction de la disponibilité d'insectes proies), la perte de bons sites de nidification ou d'alimentation (Hames et al., 2002), un accroissement chez les oiseaux de la vigilance et du temps passé à couver les œufs, et une augmentation du risque de prédation (Brotons et al., 1998). L'acidification des forêts contribue aussi au lessivage du calcium des sols, phénomène particulièrement marqué dans le nord-est du continent (Driscoll et al., 2001), où les sols ont un faible pouvoir tampon du fait de leur pH bas, et où une saturation en azote est observée (d'où un lessivage accru du calcium dû aux nitrates) (U.S. Environmental Protection Agency, 2014). Les passereaux doivent obtenir du calcium par leur alimentation durant la période de ponte (Hames et al., 2002), et une carence en calcium durant cette période peut rendre les coquilles des œufs plus mince, plus faible et plus poreuse, ce qui peut faire échouer la reproduction. Bien qu'il n'en existe pas de preuves directes, l'acidification de l'habitat de reproduction de la Paruline du Canada pourrait avoir un impact négatif sur l'espèce. L'acidification a été associée à des baisses d'effectifs chez la Grive des bois (Hames et al., 2002), ainsi que chez des passereaux d'Europe du Nord nichant dans des zones acidifiées de l'aire de reproduction de ces espèces (Graveland et Drent, 1997; Mänd et al., 2000).
5. Objectifs de population et de répartition
L'objectif de population à court terme est d'arrêter le déclin national d'ici 2025 (c'est à dire dans les 10 ans suivant la publication du présent programme de rétablissement dans le Registre public des espèces en péril), en veillant à ce que la population ne diminue pas de plus de 10 % durant cette période. L'objectif de population à long terme (après 2025) est d'assurer une tendance démographique positive sur 10 ans à l'échelle du pays.
L'objectif de répartition est de maintenir la zone d'occurrence actuelle (zone englobant la répartition géographique de l'ensemble des populations connues) au Canada.
Les objectifs de population visent à s'attaquer au déclin à long terme de l'espèce, qui a justifié la désignation de la Paruline du Canada comme espèce « menacée » (COSEPAC, 2008). Il est reconnu que l'ensemble de données du Relevé des oiseaux nicheurs (BBS, pour Breeding Bird Survey) (voir la section 3.2 – Population et répartition) comporte des lacunes en ce qui concerne la Paruline du Canada; c'est pourquoi le programme de rétablissement comprend des approches pour améliorer le suivi de l'espèce. À mesure que de nouveaux renseignements deviendront disponibles, les objectifs de population et de répartition pourront être révisés, au besoin, aux fins du rétablissement de l'espèce.
On a jugé qu'une période de 10 ans convenait pour évaluer l'évolution de la population de la Paruline du Canada. Cet espace de temps a été retenu parce qu'arrêter le déclin d'une espèce est difficile et ne peut s'accomplir en quelques années, et parce que les évaluations d'espèces du COSEPAC sont effectuées tous les 10 ans. Les critères d'évaluation du COSEPAC comprennent un examen des changements d'effectifs sur des périodes de 10 ans.
Ces objectifs seront revus au moment d'établir le rapport requis (art. 46 de la LEP) sur la mise en œuvre du présent programme de rétablissement et sur les progrès réalisés pour en atteindre les objectifs cinq ans après la publication du programme.
6. Stratégies et approches générales recommandés pour l'atteinte des objectifs
6.1 Mesures déjà achevées ou en cours
De nombreuses activités ont été lancées depuis la dernière évaluation du Comité sur la situation des espèces en péril au Canada (COSEPAC, 2008). Voici des exemples, non exhaustifs, qui illustrent les principaux domaines visés, pour mettre en contexte les grandes stratégies de rétablissement décrites à la section 6.2.
- Lancement de l'Initiative de conservation visant la Paruline du Canada, dirigée par Environnement Canada (Études d'oiseaux Canada, 2013).
- Projets dans le cadre du Programme d'intendance de l'habitat (PIH) en vue de déterminer l'habitat important en Alberta (Ball et al., 2013).
- Études observationnelles de l'utilisation de l'habitat afin de répertorier les caractéristiques qui distinguent l'habitat convenable en Colombie-Britannique (Manning Cooper and Associates Ltd., 2007), en Alberta (Krikun, 2012), en Nouvelle-Écosse (Ryan, 2012; A. Westwood, comm. pers.) et dans l'ensemble du Canada (Cumming et al., 2010).
- Études de modélisation visant à déterminer les caractéristiques importantes de l'habitat de la Paruline du Canada à diverses échelles spatiales dans tout le pays (Haché et al., 2014).
- Études observationnelles du comportement de chant afin d'améliorer les méthodes d'inventaire (Cummings, 2011; Demko, 2012) .
- Achèvement et publication des plans régionaux de conservation des oiseaux pour le Canada, qui précisent les objectifs et les mesures de conservation pour les espèces d'oiseaux prioritaires (y compris la Paruline du Canada) (Environnement Canada, 2014a).
- Études portant sur les variables explicatives à grande échelle du déclin des insectivores aériens en Amérique du Nord (voir par exemple http://homepage.usask.ca/~cam202/page11.html.
- Rapport sur la situation de la Paruline du Canada en Alberta (Ball et Bayne, en cours de révision).
- Recherche visant à déterminer les liens entre la reproduction, la migration et l'hivernage des Parulines du Canada nichant dans l'ensemble de l'aire de reproduction de l'espèce (K. Fraser, comm. pers.; S. Haché, comm. pers.).
- Suivi acoustique des migrateurs nocturnes (y compris la Paruline du Canada) près du lac Érié (Sanders et Mennill, 2014b, a).
- Rapport portant sur les connaissances actuelles concernant l'aire d'hivernage et l'écologie de la Paruline du Canada en Amérique du Sud (BirdLife International, 2014).
- Fatal Light Awareness Program (FLAP) : programme axé sur la surveillance et la prévention des collisions des oiseaux migrateurs contre des fenêtres (FLAP Canada, 2014).
- Des pratiques et initiatives d'exploitation forestière et de sylviculture dans diverses zones du pays visent à préserver les caractéristiques d'habitat considérées comme importantes pour la Paruline du Canada, ou à repérer les milieux occupés par l'espèce.
- Le ministère de la Défense nationale a effectué des activités de relevé et de suivi, et a mis en œuvre des mesures de protection de la Paruline du Canada à ses établissements dans l'ensemble du Canada.
- Articles et sites Web pour susciter l'intérêt à l'égard des mesures de rétablissement et favoriser la participation aux activités (organismes fédéraux et provinciaux et organisations non gouvernementales, comme le Service canadien de la faune, Études d'oiseaux Canada, Georgian Bay Biosphere Reserve Inc, Ontario Nature, Fatal Light Awareness Program). Exemples :
- Bird Studies Canada (document en format portable, anglais seulement)
- Ontario Nature (anglais seulement)
- borealbirds.org
- Plusieurs projets axés sur la conservation mis en œuvre au Canada et aux États-Unis incluent la Paruline du Canada dans leurs activités, notamment :
- The Boreal Avian Modeling Project (anglais seulement)
- The Songbird Initiative (anglais seulement)
- Initiative boréale canadienne
- Breeding Bird Survey, (en anglais seulement) et Relevé des oiseaux nicheurs
- Atlas des oiseaux nicheurs dans tout le Canada, et formulaires associés de signalement d'espèces rares.
6.2 Orientation stratégique pour le rétablissement
| Menace ou facteur limitatif | Stratégie générale de rétablissement | Priorité | Description générale des approches de recherche et de gestion |
|---|---|---|---|
| Perte et dégradation d'habitat, modification des systèmes naturels, espèces envahissantes et problématiques, pollution | Conservation et gestion de l'espèce et de son habitat | Élevée |
|
| Perte et dégradation d'habitat, modification des systèmes naturels, espèces envahissantes et problématiques, pollution | Conservation et gestion de l'espèce et de son habitat | Faible |
|
| Lacunes dans les connaissances | Suivi et recherche | Élevée |
|
| Lacunes dans les connaissances | Suivi et recherche | Faible |
|
| Toutes les menaces | Lois et politiques | Élevée |
|
| Toutes les menaces | Lois et politiques | Moyenne |
|
| Toutes les menaces | Éducation et sensibilisation, intendance, et partenariats | Élevée |
|
| Toutes les menaces | Éducation et sensibilisation, intendance, et partenariats | Moyenne |
|
6.3 Commentaires à l'appui du tableau de planification du rétablissement
Le rétablissement de la Paruline du Canada exigera l'engagement, la collaboration et la coopération des instances internationales, fédérales, provinciales et territoriales, des conseils de gestion des ressources fauniques, des peuples autochtones, des collectivités locales, des propriétaires fonciers, du secteur industriel et d'autres intéressés. Vu la grande aire de répartition de l'espèce, il sera important d'effectuer un suivi de l'état de l'habitat, la tendance des effectifs et la répartition de l'espèce de sorte que l'efficacité des efforts de rétablissement puisse être évaluée et améliorée, au besoin.
Conservation et gestion de l'espèce et de son habitat
On ne sait pas, à l'heure actuelle, si l'habitat de reproduction est un facteur limitatif au Canada. Néanmoins, suffisamment d'habitat de reproduction de qualité doit être géré et protégé afin d'assurer la survie de l'espèce, en particulier dans les régions où de grandes portions d'habitat pourraient être transformées ou dégradées. De plus, il faut mieux comprendre les tendances de la dynamique des populations de proies pour savoir si maintenir, améliorer ou rétablir les milieux producteurs d'insectes serait notablement profitable aux populations de Parulines du Canada.
Trouver comment déterminer l'importance relative de l'habitat de reproduction et de l'habitat autre que l'habitat de reproduction est important pour le rétablissement de l'espèce. Il est important pour assurer une conservation efficace de l'habitat et de l'espèce de savoir où apporter un soutien économique et où concentrer les activités de recherche et de suivi.
La meilleure gestion de l'habitat de reproduction n'arrivera pas à rétablir l'espèce à moins que l'habitat de migration et l'habitat d'hivernage ne soient aussi maintenus. La collaboration avec les instances internationales et les organisations non gouvernementales pour préserver, restaurer et améliorer l'habitat de migration et l'habitat d'hivernage est donc un élément tout aussi important du programme de rétablissement. Une telle collaboration devrait avoir un effet additif sur d'autres espèces en péril dont les aires d'hivernage et les aires de migration chevauchent celles de la Paruline du Canada (voir l'annexe A).
Suivi et recherche
On ne sait pas dans quelle mesure les menaces s'exerçant au pays participent à la diminution de l'effectif de la Paruline du Canada ni si les principaux facteurs responsables de cette diminution se trouvent ailleurs et agissent à un autre moment du cycle annuel de l'espèce. Il faudra adopter une approche systématique de recherche et de suivi (couvrant tous les stades du cycle biologique annuel de l'espèce et l'ensemble de la zone d'occupation) pour mieux comprendre la situation de l'espèce ainsi que les menaces et les facteurs limitatifs existant au Canada et ailleurs (voir l'annexe B pour des exemples de besoins en matière de recherche en fonction des menaces). À l'heure actuelle, le suivi de l'espèce se limite essentiellement à la partie sud de l'aire de reproduction (surtout dans l'est). Il importe donc de consacrer davantage d'attention et d'efforts à la partie nord.
Comme on vise à arrêter la baisse d'effectif de l'espèce et, ultimement, à accroître l'effectif, il convient de déterminer l'habitat qui conviendrait à l'espèce, mais que celle ci n'occupe pas actuellement. Il est important aussi de déterminer les voies, les haltes et la connectivité migratoires. Le fait de déterminer les principaux paramètres démographiques (p. ex. survie et succès de reproduction dans divers types de milieux) durant le cycle annuel de la Paruline du Canada fournira des renseignements sur les caractéristiques de l'habitat qui convient le mieux ainsi que sur les activités pertinentes ou les localités préoccupantes pour l'espèce, la taille des populations, etc. Il sera alors nécessaire de bâtir sur ces bases et de valider des modèles d'habitat à l'échelle nationale et régionale pour mieux comprendre à quel endroit l'espèce pourrait se reproduire, et contribuer aux efforts de protection de l'habitat. Dans l'aire d'hivernage, les programmes de suivi sont moins nombreux, mais ils sont essentiels. Il faut donc les développer et les mettre en œuvre pour obtenir de meilleures informations sur l'utilisation de l'habitat et sur les tendances locales des effectifs et de l'habitat.
Si certaines recherches ont précisé les besoins en matière d'habitat et les facteurs limitatifs de la Paruline du Canada depuis la rédaction du rapport de situation, en 2008, la plus grande partie de ces travaux a été menée à seulement quelques sites d'étude dans l'est des États-Unis (Vermont et New Hampshire) (Reitsma et al., 2010). Il faudrait étendre les recherches dans la partie canadienne de l'aire de reproduction de l'espèce, et les axer sur les objectifs suivants : 1) déterminer l'importance relative des menaces évoquées; 2) déterminer les caractéristiques d'habitat qui maximisent les effectifs et le succès de reproduction; 3) établir si les données d'effectif fournissent une mesure valable de la qualité de l'habitat.
Tandis que les activités de suivi et de recherche nécessaires sont menées, les connaissances scientifiques courantes peuvent constituer une base permettant de protéger les habitats connus et d'atténuer les menaces pesant sur l'espèce.
Lois et politiques
Il existe de multiples moyens législatifs et volontaires pour protéger la Paruline du Canada et son habitat au Canada.
Les interdictions générales prévues par la Loi de 1994 sur la convention concernant les oiseaux migrateurs et ses règlements protègent les nids et les œufs de la Paruline du Canada, peu importe où ils se trouvent au Canada et quelle que soit la propriété des terres. Les nids et les œufs peuvent néanmoins être perturbés ou détruits par mégarde au cours de nombreuses activités, notamment la coupe d'arbres. Durant la période de reproduction, il faut éviter de mener toute activité potentiellement destructrice ou perturbatrice aux localités où la Paruline du Canada est susceptible d'être observée ou aux localités où sa présence est connue (Environment Canada, 2014). Différentes avenues, comme les politiques et règlements d'aménagement du territoire et les évaluations environnementales, peuvent aussi contribuer à assurer ce type de protection.
Il faut élaborer et mettre en œuvre des pratiques de gestion bénéfiques (PGB) et des politiques connexes pour la Paruline du Canada, ses proies et leur habitat, et ces PGB et politiques doivent être fondées sur la meilleure information scientifique disponible. Les PGB et les politiques connexes peuvent viser diverses menaces connues et présumées, y compris l'exploitation forestière et la sylviculture, l'exploration et l'exploitation de ressources énergétiques et minières, les collisions contre les fenêtres, le broutage et le pâturage et les espèces problématiques. Les PGB pour la Paruline du Canada doivent être intégrées à celles qui visent d'autres espèces afin de préserver les paysages hétérogènes qui constituent une mosaïque dynamique de conditions d'habitat dont profiteront plusieurs espèces. Chaque fois que cela est possible, il faut appliquer une approche plurispécifique en matière de rétablissement. Les PGB qui seront adoptées par les gouvernements, le secteur industriel et même les particuliers pourront jouer un rôle important dans les efforts en cours dans l'ensemble de l'aire de répartition de l'espèce, et seront nécessaires pour favoriser le rétablissement et la conservation de la Paruline du Canada sur une vaste échelle, d'un bout à l'autre du continent et jusqu'en Amérique du Sud.
Les normes et codes d'application volontaire adoptés par le secteur privé (p. ex. la certification de l'aménagement forestier durable par des tiers, les systèmes de cotation internationaux qui reconnaissent l'excellence des bâtiments écologiques) peuvent aider à réduire certaines menaces qui pèsent sur l'espèce et ses proies.
Par-delà les frontières du Canada, la collaboration internationale en matière de programmes de recherche et d'intendance sera importante pour assurer la conservation et le rétablissement de la Paruline du Canada, vu les menaces qui peuvent peser sur l'espèce dans les aires autres que les aires de reproduction. Une collaboration du genre a déjà été établie par l'Initiative de conservation visant la Paruline du Canada, qui mènera des activités à l'échelle internationale dans les années à venir.
Éducation et sensibilisation, intendance, et partenariats
Il faudrait favoriser les collaborations avec divers ordres de gouvernement, des propriétaires fonciers, des forestiers, le secteur industriel et des propriétaires d'animaux domestiques (entre autres). Il faut réaliser des activités d'intendance à des endroits stratégiques dans l'ensemble de l'aire de répartition de la Paruline du Canada au pays, en particulier là où il y a un risque accru de développement dans un proche avenir. À ces endroits où le développement est prévisible, le besoin de PGB et de politiques pertinentes se fera grandissant. Il faut encourager la conformité aux lois et règlements, aux politiques et aux PGB qui protègent l'espèce. Les travaux d'exploration de ressources énergétiques et minières s'intensifient dans certaines parties de la région boréale du Canada, où les menaces pour l'espèce ne feront qu'augmenter avec le temps.
Préserver et rehausser l'habitat de reproduction de la Paruline du Canada exigera de favoriser la conservation et l'intendance à une vaste échelle. Les principales mesures à encourager comprennent les pratiques d'exploitation forestière et les pratiques sylvicoles qui fournissent de l'habitat de reproduction et réduisent le risque de perturbation ou de destruction des nids et des œufs, ainsi que les pratiques qui diminuent le risque de collisions contre des fenêtres. De plus, parce que la grande étendue de l'aire de répartition de l'espèce et la relative inaccessibilité de certaines parties de cette aire (p. ex. la partie nord) compliquent le suivi et les relevés à grande échelle, il est crucial d'élaborer des relevés ciblés et des méthodes de relevés novatrices (p. ex. le suivi acoustique), et de promouvoir la participation bénévole. Il convient notamment d'intéresser les clubs d'ornithologues qui connaissent les lieux où il y a de fortes densités de nicheurs et les scientifiques amateurs qui participent aux programmes d'atlas d'oiseaux et de relevés d'oiseaux nicheurs. De plus, il faut continuer à favoriser la collecte de données par les citoyens (p. ex. eBird) pour bonifier les activités de recherche et de suivi en cours.
Les normes et codes d'application volontaire adoptés par le secteur privé, comme la certification de l'aménagement forestier durable par des tiers, les systèmes de cotation internationaux qui reconnaissent l'excellence des bâtiments écologiques, et divers programmes, comme le Programme d'accréditation de la lutte antiparasitaire intégrée, peuvent aussi aider à réduire certaines menaces qui pèsent sur l'espèce et ses proies.
7. Habitat essentiel
Selon l'alinéa 41(1)c) de la Loi sur les espèces en péril, le programme de rétablissement doit comporter la désignation de l'habitat essentiel de l'espèce dans la mesure du possible.
7.1 Désignation de l'habitat essentiel de l'espèce
L'examen de l'aire de répartition géographique de l'espèce, de la spécificité de son habitat, de la taille de sa population et des menaces qui pèsent sur elle indique que l'habitat essentiel devrait être désigné à l'échelle du paysage Note de bas de page3. Si on comprend de façon générale le caractère convenable de l'habitat (voir la section 3.3) et qu'une certaine modélisation du caractère convenable de l'habitat a été réalisée (Haché et al., 2014), il reste à déterminer si l'habitat est limitatif au Canada. Les renseignements dont on dispose ne permettent pas de désigner l'habitat essentiel à l'échelle du paysage pour les raisons suivantes :
- On comprend mal quelle est la configuration appropriée des caractéristiques biophysiques importantes pour l'espèce à l'échelle du paysage, et les données manquent à cet égard.
- Les besoins en matière d'habitat peuvent varier dans l'aire de répartition de l'espèce. Les unités de gestion (c.-à-d. les unités géographiques au sein desquelles l'habitat essentiel serait géré) doivent être identifiées de manière à tenir compte le mieux possible de la variation de l'utilisation de l'habitat et des modes de gestion.
- Il y a un manque de données sur la présence et l'abondance de la Paruline du Canada dans de grandes portions de son aire de répartition. Sans ces données, les modèles servant à prévoir l'habitat essentiel qui sont fondés sur les renseignements dont on dispose actuellement auront des capacités limitées pour ces portions.
- Dans le cas de la Paruline du Canada, on ne sait pas exactement si certains habitats présentant des caractéristiques biophysiques spécifiques conviennent mieux à l'espèce que d'autres sur le plan fonctionnel. Par exemple, certains habitats pourraient abriter de plus fortes densités d'individus ou de couples, et/ou résulter en un succès de reproduction plus élevé.
- On connaît peu les relations entre les perturbations causées par l'activité humaine et la qualité de l'habitat. Il est nécessaire de mieux comprendre ces relations pour pouvoir offrir à la Paruline du Canada une quantité suffisante d'habitat convenable et déterminer l'échelle et l'intensité auxquelles les activités pourraient probablement entraîner la destruction de l'habitat essentiel.
Un calendrier des études a été élaboré en vue d'obtenir les renseignements nécessaires à la désignation de l'habitat essentiel qui suffira pour atteindre les objectifs en matière de population et de répartition. La désignation de l'habitat essentiel sera présentée, soit dans une mise à jour du programme de rétablissement, soit dans un plan d'action.
7.2 Calendrier des études visant à désigner l'habitat essentiel
Afin d'orienter le calendrier des études, le groupe du Projet de modélisation de l'avifaune boréale a entrepris récemment une modélisation qui aidera à comprendre les caractéristiques de l'utilisation de l'habitat par la Paruline du Canada (Haché et al., 2014). Ces auteurs ont évalué les caractéristiques de l'habitat de la Paruline du Canada dans tout le pays d'après des données de points d'écoute, des paramètres de classification des terres (couverture terrestre, perturbation et topographie) et des données environnementales (climat). Si les résultats ont fait mieux connaître l'utilisation que fait l'espèce de l'habitat au Canada, ils ne peuvent servir à désigner l'habitat qui est essentiel à la survie ou au rétablissement de l'espèce, en raison du manque de renseignements pertinents (comme il est exposé à la section 7.1).
Voici le calendrier des études qui permettront de désigner l'habitat essentiel.
| Description de l'activité | Justification | Échéancier |
|---|---|---|
| Déterminer les unités de gestion qui conviennent d'après les besoins de l'espèce en matière d'habitat dans l'ensemble de son aire de répartition. | Les besoins en matière d'habitat peuvent varier dans l'aire de répartition de l'espèce. Il convient de définir des unités de gestion afin de tenir compte le mieux possible de la variation de l'utilisation de l'habitat. | 2016 |
| Augmenter le suivi à des endroits stratégiques. | Les données sur les effectifs, la productivité et d'autres mesures de la qualité de l'habitat sont rares pour de nombreuses régions du pays. Il faut augmenter le suivi à des endroits choisis pour valider et améliorer les récents modèles d'habitat (Haché et al., 2014). | 2016-2020 |
| Établir la configuration appropriée des caractéristiques biophysiques à l'échelle du paysage. | Pour désigner l'habitat essentiel à l'échelle du paysage, il est nécessaire de comprendre les caractéristiques biophysiques que requiert l'espèce à cette échelle, et de déterminer comment ces caractéristiques doivent être configurées pour répondre aux besoins de l'espèce. | 2016-2019 |
| Déterminer la qualité de l'habitat pour l'ensemble des unités de gestion et au sein de chacune d'entre elles. | Les données sur les effectifs, la productivité et d'autres mesures de la qualité de l'habitat peuvent permettre de repérer des zones particulières qui contribuent fortement à la survie ou au rétablissement de la Paruline du Canada. | 2016-2020 |
| Déterminer les seuils d'étendue et d'intensité auxquels les activités humaines deviendraient susceptibles de détruire l'habitat qui convient à l'espèce. | Il est nécessaire de mieux comprendre les relations entre les perturbations causées par l'activité humaine et la qualité de l'habitat pour pouvoir offrir à la Paruline du Canada une quantité suffisante d'habitat convenable et déterminer les seuils d'étendue et d'intensité à partir desquels les activités humaines deviendraient susceptibles de détruire l'habitat essentiel. | 2016-2020 |
| Déterminer la quantité d'habitat convenable requise pour atteindre les objectifs de population et de répartition. | On ne sait pas si l'habitat constitue un facteur limitatif pour la Paruline du Canada au pays. Il faut évaluer si l'habitat au pays est suffisant pour atteindre les objectifs de population et de répartition. | 2020 |
| Élaborer et valider des modèles d'habitat pour déterminer les endroits où les caractéristiques biophysiques existent en quantité, en qualité et en configuration voulues dans chaque unité de gestion afin d'atteindre les objectifs de population et de répartition. | Les résultats des études susmentionnées permettront de bâtir des modèles permettant de déterminer la localisation géographique, la quantité et la qualité de l'habitat qui devrait être désigné comme habitat essentiel pour la Paruline du Canada. | 2021 |
8. Mesure des progrès
Les indicateurs de rendement présentés plus bas permettront d'évaluer les progrès accomplis vers l'atteinte des objectifs de population et de répartition.
- À court terme (10 ans; avant 2025), les tendances à la baisse des populations sont stoppées ou inversées, de sorte que les populations canadiennes de Parulines du Canada ne perdent pas plus de 10 % de leur effectif durant cette période.
- À long terme (après 2025), une tendance positive sur 10 ans est obtenue (l'effectif augmente).
- La zone d'occurrence de la reproduction de la Paruline du Canada est préservée dans tout le pays.
9. Énoncé sur les plans d'action
Un ou plusieurs plans d'action visant la Paruline du Canada seront publiés dans le Registre public des espèces en péril dans les cinq ans suivant la publication du programme de rétablissement.
10. Références
- Angers, V.A., C. Messier, M. Beaudet et A. Leduc. 2005. Comparing composition and structure in old-growth and harvested (selection and diameter-limit cuts) northern hardwood stands in Quebec. Forest Ecology and Management 217(2):275 293.
- Bakermans, M.H., A.D. Rodewald, A.C. Vitz et C. Rengifo. 2012. Migratory bird use of shade coffee: the role of structural and floristic features. Agroforestry Systems 85(1):85 94.
- Bakermans, M.H., A.C. Vitz, A.D. Rodewald et C.G. Rengifo. 2009. Migratory songbird use of shade coffee in the Venezuelan Andes with implications for conservation of Cerulean Warbler. Biological Conservation 142(11):2476 2483.
- Ball, J.R., et E. Bayne. En cours de révision. Status of the Canada Warbler (Cardellina canadensis) in Alberta. Alberta Environment and Sustainable Resource Development, Edmonton, AB.
- Ball, J.R., P. Sólymos, E. Bayne, T. Habib, D. Stepnisky, L. Mahon, F. Schmiegelow, S.J. Song et S. Cumming. 2013. Determination of habitat associations and development of best management practices of Canada Warblers in mixed-wood boreal forests in Alberta. Rapport inédit présenté au Programme d'intendance de l'habitat des espèces en péril, Environnement Canada, Edmonton, AB.
- Baudvin, H. 1997. Barn Owl (Tyto alba) and Long-eared Owl (Asio otus) mortality along motorways in Bourgogne-Champagne: report and suggestions, p. 58 61 in Biology and Conservation of Owls of the Northern Hemisphere: 2nd International Symposium. U.S. Dept. of Agriculture, Forest Service, North Central Forest Experiment Station, St. Paul, MN.
- BDNF. 2014. Base de données nationale sur les forêts. Ministère des Ressources naturelles Canada, Service canadien des forêts, Ottawa, ON. (consulté le 20 août 2014].
- Bent, A.C. 1953. Life Histories of North American Wood Warblers: Order Passeriformes. US Government Printing Office, Washington D.C.
- Benton, T.G., D.M. Bryant, L. Cole et H.Q. Crick. 2002. Linking agricultural practice to insect and bird populations: a historical study over three decades. Journal of Applied Ecology 39(4):673 687.
- Bergeron, Y., et N.J. Fenton. 2012. Boreal Forests of Eastern Canada Revisited: Old growth, nonfire disturbances, forest succession, and biodiversity. Botanique 90(6):509 523 (comprend un résumé en français).
- Birch, R., et D. Kaye. 2012. Global mining finance: 2012, Toronto, ON.
- BirdLife International. 2014. Canada Warbler (Cardellina canadensis) wintering distribution and ecology: South America. BirdLife International, Quito, Équateur.
- BirdLife International et NatureServe. 2013. Bird species distribution maps of the world, BirdLife International and NatureServe, Cambridge, Royaume Uni et Arlington, VA, disponible sur demande (consulté en août 2014).
- Bishop, C.A., et J.M. Brogan. 2013. Estimates of avian mortality attributed to vehicle collisions in Canada (Estimation de la mortalité aviaire attribuable aux collisions automobiles au Canada). Avian Conservation and Ecology 8(2):2 (comprend un résumé en français).
- Blancher, P. 2013. Estimated number of birds killed by house cats (Felis catus) in Canada (Estimation du nombre d'oiseaux tués par les chats domestiques [Felis catus] au Canada). Avian Conservation and Ecology 8(2):3 (comprend un résumé en français).
- Blancher, P.J., et D.K. McNicol. 1991. Tree swallow diet in relation to wetland acidity. Revue canadienne de zoologie 69(10):2629 2637 (comprend un résumé en français).
- Blancher, P.J., K.V. Rosenberg, A.O. Panjabi, B. Altman, J. Bart, C.J. Beardmore, G.S. Butcher, D. Demarest, R. Dettmers, E.H. Dunn, W. Easton, W.C. Hunter, E.E. Iñigo Elias, D.N. Pashley, C.J. Ralph, C. Rich, C.M. Rustay, J.M. Ruth et T. Will. 2007. Guide to the Partners in Flight Population Estimates Database, version: North American Landbird Conservation Plan 2004, Partners in Flight Technical Series No. 5.
- Boatman, N.D., N.W. Brickle, J.D. Hart, T.P. Milsom, A.J. Morris, A.W. Murray, K.A. Murray et P.A. Robertson. 2004. Evidence for the indirect effects of pesticides on farmland birds. Ibis 146(s2):131 143.
- Boreal Songbird Initiative. 2012. Boreal bird declines and human disturbances fact sheet. (consulté le 20 août 2014).
- Both, C., S. Bouwhuis, C. Lessells et M.E. Visser. 2006. Climate change and population declines in a long-distance migratory bird. Nature 441(7089):81 83.
- Both, C., C.A. Van Turnhout, R.G. Bijlsma, H. Siepel, A.J. Van Strien et R.P. Foppen. 2009. Avian population consequences of climate change are most severe for long-distance migrants in seasonal habitats. Proceedings of the Royal Society B: Biological Sciences 277:1259 1266.
- Brooks, D.R., J.E. Bater, S.J. Clark, D.T. Monteith, C. Andrews, S.J. Corbett, D.A. Beaumont et J.W. Chapman. 2012. Large carabid beetle declines in a United Kingdom monitoring network increases evidence for a widespread loss in insect biodiversity. Journal of Applied Ecology 49(5):1009 1019.
- Brotons, L., M. Magrans, L. Ferrús et J. Nadal. 1998. Direct and indirect effects of pollution on the foraging behaviour of forest passerines during the breeding season. Revue canadienne de zoologie 76(3):556 565 (comprend un résumé en français).
- Calvert, A.M., C.A. Bishop, R.D. Elliot, E.A. Krebs, T.M. Kydd, C.S. Machtans et G.J. Robertson. 2013. A synthesis of human-related avian mortality in Canada (Synthèse des sources de mortalité aviaire d'origine anthropique au Canada). Avian Conservation and Ecology 8(2):11 (comprend un résumé en français).
- Campbell, R. W., M. I. Preston, M. Phinney, C. Siddle, et J. Deal. 2007. Wildlife Data Centre Featured Species - Canada Warbler. Wildlife Afield 4(1): 95-160.
- Case, R.M. 1978. Interstate highway road-killed animals: a data source for biologists, Wildlife Society Bulletin 6(1):8 13.
- Chace, J.F., S.D. Faccio et A. Chacko. 2009. Canada Warbler habitat use of northern hardwoods in Vermont. Northeastern Naturalist 16(4):491 500.
- Chamberlain, D.E., R.J. Fuller, R.G.H. Bunce, J.C. Duckworth et M. Shrubb. 2000. Changes in the abundance of farmland birds in relation to the timing of agricultural intensification in England and Wales. Journal of Applied Ecology 37(5):771 788.
- Cheng, R., et P. Lee. 2014. Canada's industrial concessions: a spatial analysis. Global Forest Watch Canada.
- Commission de coopération environnementale de l'Amérique du Nord. 2003. Le DDT n'est plus utilisé en Amérique du Nord, (Format PDF, 450 Ko). Commission de coopération environnementale de l'Amérique du Nord, Montréal, QC.
- Cooper, J., K.A. Enns et M.G. Shepard. 1997. Status of the Canada Warbler in British Columbia. 0772629978, Ministry of Environment, Lands and Parks, Victoria, BC.
- COSEPAC. 2008. Évaluation et Rapport de situation du COSEPAC sur la Paruline du Canada, (Format PDF) Wilsonia canadensis, au Canada. ISBN 9780662041368, Comité sur la situation des espèces en péril au Canada, Ottawa, ON.
- COSEPAC. 2009. Évaluation et Rapport de situation du COSEPAC sur l'Engoulevent bois-pourri, (Format PDF) Caprimulgus vociferus, au Canada. Comité sur la situation des espèces en péril au Canada, Ottawa, ON.
- COSEPAC. 2012. Évaluation et Rapport de situation du COSEPAC sur le Pioui de l'Est, (Format PDF) Contopus virens, au Canada. Comité sur la situation des espèces en péril au Canada, Ottawa, ON.
- Côté, S.D., T.P. Rooney, J. P. Tremblay, C. Dussault et D.M. Waller. 2004. Ecological impacts of deer overabundance. Annual Review of Ecology, Evolution, and Systematics 35:113 147.
- Cumming, S.G., K.L. Lefevre, E. Bayne, T. Fontaine, F.K. Schmiegelow et S.J. Song. 2010. Toward conservation of Canada's boreal forest avifauna: design and application of ecological models at continental extents (Conservation de l'avifaune de la forêt boréale au Canada : élaboration et application de modèles écologiques à l'échelle continentale). Avian Conservation and Ecology 5(2):8 (comprend un résumé en français).
- Cumming, S.G., D. Stralberg, K.L. Lefevre, P. Sólymos, E.M. Bayne, S. Fang, T. Fontaine, D. Mazerolle, F.K.A. Schmiegelow et S.J. Song. 2014. Climate and vegetation hierarchically structure patterns of songbird distribution in the Canadian boreal region. Ecography 37(2):137 151.
- Cummings, G. 2011. Using Vocal Behaviour to Reform the Conservation of Canada Warblers (Wilsonia canadensis). Mémoire de spécialisation de baccalauréat, Dalhousie University, Halifax, NS.
- Davis, S., V. Heywood, O. Herrera-Macbryde, J. Villa-Lobos et A. Hamilton. 1997. Centres of plant diversity: a guide and strategy for their conservation. Volume 3, IUCN Publications Unit, Cambridge, Royaume-Uni. (consulté en août 2014).
- de Groot, W.J., M.D. Flannigan et A.S. Cantin. 2013. Climate change impacts on future boreal fire regimes, Forest Ecology and Management 294:35 44.
- DeGraaf, R.M., W.M. Healy et R.T. Brooks. 1991. Effects of thinning and deer browsing on breeding birds in New England oak woodlands. Forest Ecology and Management 41(3):179 191.
- DeGraaf, R.M., et M. Yamasaki. 2003. Options for managing early-successional forest and shrubland bird habitats in the northeastern United States. Forest Ecology and Management 185(1):179 191.
- Demko, A. D., L. R. Reitsma et A. Staicer. 2013. Two song categories in the Canada Warbler (Cardellina canadensis). The Auk 130(4):609-616.
- Depew, D.C., N.M. Burgess et L.M. Campbell. 2013. Modelling mercury concentrations in prey fish: derivation of a national-scale common indicator of dietary mercury exposure for piscivorous fish and wildlife. Environmental Pollution 176:234 243.
- Dirzo, R., H.S. Young, M. Galetti, G. Ceballos, N.J. Isaac et B. Collen. 2014. Defaunation in the Anthropocene, Science 345(6195):401 406.
- Drapeau, P., A. Leduc, J. F. Giroux, J. P.L. Savard, Y. Bergeron et W.L. Vickery. 2000. Landscape-scale disturbances and changes in bird communities of boreal mixed wood forests, Ecological Monographs 70(3):423 444.
- Driscoll, C.T., G.B. Lawrence, A.J. Bulger, T.J. Butler, C.S. Cronan, C. Eagar, K.F. Lambert, G.E. Likens, J.L. Stoddard et K.C. Weathers. 2001. Acidic deposition in the northeastern United States: sources and inputs, ecosystem effects, and management strategies, BioScience 51(3):180 198.
- Drummond, M.A., et T.R. Loveland. 2010. Land-use pressure and a transition to forest-cover loss in the eastern United States, BioScience 60(4):286 298.
- eBird. 2014. eBird: an online database of bird distribution and abundance, version 2, Audubon and Cornell Lab of Ornithology, Ithaca, NY. Disponible à l'adresse : (consulté le 10 septembre 2014).
- Edmonds, S.T., D.C. Evers, D.A. Cristol, C. Mettke-Hofmann, L.L. Powell, A.J. McGann, J.W. Armiger, O.P. Lane, D.F. Tessler et P. Newell. 2010. Geographic and seasonal variation in mercury exposure of the declining Rusty Blackbird. The Condor 112(4):789 799.
- Edmonds, S.T., N.J. O'Driscoll, N.K. Hillier, J.L. Atwood et D.C. Evers. 2012. Factors regulating the bioavailability of methylmercury to breeding Rusty Blackbirds in northeastern wetlands. Environmental Pollution 171:148 154.
- Environment Canada. 2014. Avoidance guidelines related to incidental take of migratory birds in Canada. Environnement Canada, Gatineau, QC.
- Environnement Canada. 2011a. Présence et concentrations des pesticides prioritaires dans certains écosystèmes aquatiques canadiens, (Format PDF, 3 Mo). Environnement Canada, Direction des sciences et de la technologie de l'eau, Gatineau, QC.
- Environnement Canada. 2011b. Situation des oiseaux au Canada. Environnement Canada, Gatineau, QC.
- Environment Canada. 2013. Loi sur les espèces en péril – directives opérationnelles aux praticiens du rétablissement. Environnement Canada, Service canadien de la faune, Ottawa, ON.
- Environnement Canada. 2014a. Régions de conservation des oiseaux et Stratégies de conservation. Environnement Canada, Ottawa, ON. (consulté le 19 septembre 2014).
- Environnement Canada. 2014b. Inventaire national des rejets de polluants. Environnement Canada, Gatineau, QC. (consulté le 4 septembre 2014).
- Environnement Canada. 2014c. Résultats du Relevé des oiseaux nicheurs. Environnement Canada, Gatineau, QC. (consulté le 15 août 2014).
- Etter, A., C. McAlpine, K. Wilson, S. Phinn et H. Possingham. 2006. Regional patterns of agricultural land use and deforestation in Colombia. Agriculture, Ecosystems & Environment 114(2):369 386.
- Études d'oiseaux Canada. 2013. Initiative de conservation visant la Paruline du Canada, (consulté le 19 septembre 2014; en anglais et en français). Études d'oiseaux Canada, Port Rowan, ON.
- Evers, D.C., K.A. Williams, M.W. Meyer, A.M. Scheuhammer, N. Schoch, A.T. Gilbert, L. Siegel, R.J. Taylor, R. Poppenga et C.R. Perkins. 2011. Spatial gradients of methylmercury for breeding common loons in the Laurentian Great Lakes region. Ecotoxicology 20(7):1609 1625.
- Fitzgerald, W.F., D.R. Engstrom, R.P. Mason et E.A. Nater. 1998. The case for atmospheric mercury contamination in remote areas. Environmental Science & Technology 32(1):1 7.
- Flannigan, M., B. Stocks, M. Turetsky et M. Wotton. 2009. Impacts of climate change on fire activity and fire management in the circumboreal forest. Global Change Biology 15(3):549 560.
- FLAP Canada. 2014. Fatal Light Awareness Program, FLAP Canada, Toronto, ON, disponible (consulté le 19 septembre 2014).
- Flockhart, D.T. 2010. Timing of events on the breeding grounds for five species of sympatric warblers. Journal of Field Ornithology 81(4):373 382.
- Foster, G.N. 1991. Conserving insects of aquatic and wetland habitats, with special reference to beetles, p. 237 262, et al.,in The Conservation of Insects and their Habitats, 15th Symposium of the Royal Entomological Society of London. Academic Press, Londres, Royaume-Uni.
- Freedman, B. 1995. Environmental ecology: the ecological effects of pollution, disturbance, and other stresses. Academic Press, San Diego (Californie), 606 p.
- Friend, M., et J.C. Franson. 1999. Field manual of wildlife diseases: general field procedures and diseases of birds. US Geological Survey, Biological Resources Division Information and Technology Report 1999-2001, DTIC Document.
- Gard, N.W., M.J. Hooper et R.S. Bennett. 1993. Effects of pesticides and contaminants on neotropical migrants, p. 310 314 in D.M. Finch et P.W. Strange (éd.), Status and Management of Neotropical Migratory Birds, Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station, Fort Collins, CO.
- Gibbons, D., C. Morrissey et P. Mineau. 2014. A review of the direct and indirect effects of neonicotinoids and fipronil on vertebrate wildlife. Environmental Science and Pollution Research:1 16.
- Girardin, M.P., A.A. Ali, C. Carcaillet, O. Blarquez, C. Hély, A. Terrier, A. Genries et Y. Bergeron. 2013. Vegetation limits the impact of a warm climate on boreal wildfires. New Phytologist 199(4):1001 1011.
- Goodnow, M., et L. Reitsma. 2011. Nest-site selection in the Canada Warbler (Wilsonia canadensis) in central New Hampshire. Revue canadienne de zoologie 89(12):1172 1177 (comprend un résumé en français).
- Goulson, D. 2014. Pesticides linked to bird declines. Nature 511:295 296.
- Gouvernement du Canada. 2009. Politiques de la Loi sur les espèces en péril : cadre général de politiques [ébauche], Loi sur les espèces en péril. Séries de politiques et de lignes directrices, Environnement Canada, Ottawa, ON.
- Graveland, J. 1998. Effects of acid rain on bird populations. Dossiers environnement 6(1):41 54 (comprend un résumé en français).
- Graveland, J., et R. Drent. 1997. Calcium availability limits breeding success of passerines on poor soils. Journal of Animal Ecology 66(2):279 288.
- Greenberg, R., et S.M. Matsuoka. 2010. Special section: rangewide ecology of the declining Rusty Blackbird - Rusty Blackbird: Mysteries of a Species in Decline. The Condor 112(4):770 777.
- Haché, S., P. Solymos, T. Fontaine, E. Bayne, S. Cumming, F. Schmiegelow et D. Stralberg. 2014. Habitat of Olive-sided Flycatcher, Canada Warbler, and Common Nighthawk in Canada. Boreal Avian Modelling Project, Edmonton, AB.
- Hagen, J.M., P.S. McKinley, A.L. Meehan et S.L. Grove. 1997. Diversity and abundance of landbirds in a northeastern industrial forest. Journal of Wildlife Management 61:718 735.
- Hallmann, C.A., R.P. Foppen, C.A. van Turnhout, H. de Kroon et E. Jongejans. 2014. Declines in insectivorous birds are associated with high neonicotinoid concentrations. Nature 511:341 343.
- Hallworth, M., P.M. Benham, J.D. Lambert et L. Reitsma. 2008a. Canada Warbler (Wilsonia canadensis) breeding ecology in young forest stands compared to a Red maple (Acer rubrum) swamp. Forest Ecology and Management 255(3):1353 1358.
- Hallworth, M., A. Ueland, E. Anderson, J.D. Lambert et L. Reitsma. 2008b. Habitat selection and site fidelity of Canada Warblers (Wilsonia canadensis) in central New Hampshire. The Auk 125(4):880 888.
- Hames, R.S., K.V. Rosenberg, J.D. Lowe, S.E. Barker et A.A. Dhondt. 2002. Adverse effects of acid rain on the distribution of the Wood Thrush Hylocichla mustelina in North America. Proceedings of the National Academy of Sciences 99(17):11235 11240.
- Hansen, M., P. Potapov, R. Moore, M. Hancher, S. Turubanova, A. Tyukavina, D. Thau, S. Stehman, S. Goetz et T. Loveland. 2013. High-resolution global maps of 21st century forest cover change. Science 342(6160):850 853.
- Hawley, D.M., K.K. Hallinger et D.A. Cristol. 2009. Compromised immune competence in free-living Tree Swallows exposed to mercury. Ecotoxicology 18(5):499 503.
- Henderson, A., S.P. Churchill et J.L. Luteyn. 1991. Neotropical plant diversity. Nature 351(6321):21 22.
- Hladik, M.L., D.W. Kolpin et K.M. Kuivila. 2014. Widespread occurrence of neonicotinoid insecticides in streams in a high corn and soybean producing region, USA. Environmental Pollution 193:189 196.
- Hobson, K., et E. Bayne. 2000a. The effects of stand age on avian communities in aspen-dominated forests of central Saskatchewan, Canada. Forest Ecology and Management 136(1):121 134.
- Hobson, K.A., et E. Bayne. 2000b. Breeding bird communities in boreal forest of western Canada: consequences of "unmixing" the mixedwoods. The Condor 102(4):759 769.
- Hobson, K.A., et E. Bayne. 2000c. Effects of forest fragmentation by agriculture on avian communities in the southern boreal mixedwoods of western Canada. The Wilson Bulletin 112(3):373 387.
- Hobson, K.A., E.M. Bayne et S.L. Van Wilgenburg. 2002. Large-scale conversion of forest to agriculture in the boreal plains of Saskatchewan. Conservation Biology 16(6):1530 1541.
- Hobson, K.A., et J. Schieck. 1999. Changes in bird communities in boreal mixedwood forest: harvest and wildfire effects over 30 years. Ecological Applications 9(3):849 863.
- Hobson, K.A., A.G. Wilson, S.L. Van Wilgenburg et E.M. Bayne. 2013. An estimate of nest loss in Canada due to industrial forestry operations (Estimation de la perte de nids attribuable aux activités forestières industrielles au Canada). Avian Conservation and Ecology 8(2):5 (comprend un résumé en français).
- Holmes, S.B. 1998. Reproduction and nest behaviour of Tennessee Warblers Vermivora peregrina in forests treated with Lepidoptera specific insecticides. Journal of Applied Ecology 35(2):185 194.
- Holt, C.A., R.J. Fuller et P.M. Dolman. 2013. Deer reduce habitat quality for a woodland songbird: evidence from settlement patterns, demographic parameters, and body condition. The Auk 130(1):13 20.
- Jones, T., et W. Cresswell. 2010. The phenology mismatch hypothesis: are declines of migrant birds linked to uneven global climate change? Journal of Animal Ecology 79(1):98 108.
- Keller, R.H., L. Xie, D.B. Buchwalter, K.E. Franzreb et T.R. Simons. 2014. Mercury bioaccumulation in Southern Appalachian birds, assessed through feather concentrations. Ecotoxicology 23(2):304 316.
- King, D.I., et R.M. DeGraaf. 2000. Bird species diversity and nesting success in mature, clearcut and shelterwood forest in northern New Hampshire, USA. Forest Ecology and Management 129(1):227 235.
- Klemens, J., R. Harper, J. Frick, A. Capparella, H. Richardson et M. Coffey. 2000. Patterns of organochlorine pesticide contamination in neotropical migrant passerines in relation to diet and winter habitat. Chemosphere 41(7):1107 1113.
- Krikun, R. 2012. Canada Warbler Project. The Warbler, été 2012:5 6.
- Krzic, M., H. Page, R. Newman et K. Broersma. 2004. Aspen regeneration, forage production, and soil compaction on harvested and grazed boreal aspen stands. BC Journal of Ecosystems and Management 5(2):30 38.
- Larsen, T.H., F. Escobar et I. Armbrecht. 2011. Insects of the tropical Andes: diversity patterns, processes and global change, pages 228-244 in S. K. Herzog, R. Martinez, P. M. Jorgensen, and H. Tiessen (eds.), Climate Change and Biodiversity in the Tropical Andes. Inter-American Institute of Global Change Research (IAI) and Scientific Committee on Problems of the Environment (SCOPE), São José dos Campos et Paris.
- Lebbin, D.J., M.J. Parr et G.H. Fenwick. 2010. The American Bird Conservancy guide to bird conservation. University of Chicago Press, Chicago, IL, 456 p.
- Longcore, T., C. Rich, P. Mineau, B. MacDonald, D.G. Bert, L.M. Sullivan, E. Mutrie, S.A. Gauthreaux Jr, M.L. Avery et R.L. Crawford. 2012. An estimate of avian mortality at communication towers in the United States and Canada. PLoS One 7(4):e34025.
- Longcore, T., C. Rich, P. Mineau, B. MacDonald, D.G. Bert, L.M. Sullivan, E. Mutrie, S.A. Gauthreaux Jr, M.L. Avery et R.L. Crawford. 2013. Avian mortality at communication towers in the United States and Canada: which species, how many, and where? Biological Conservation 158:410 419.
- Loss, S.R., T. Will, S.S. Loss et P.P. Marra. 2014a. Bird-building collisions in the United States: estimates of annual mortality and species vulnerability. The Condor 116(1):8 23.
- Loss, S.R., T. Will et P.P. Marra. 2014b. Estimation of bird-vehicle collision mortality on US roads. The Journal of Wildlife Management 78(5):763 771.
- Loss, S.R., T. Will et P.P. Marra. 2014c. Refining estimates of bird collision and electrocution mortality at power lines in the United States. PLoS One 9(7):e101565.
- Machtans, C.S., K.J. Kardynal et P.A. Smith. 2014. How well do regional or national Breeding Bird Survey data predict songbird population trends at an intact boreal site? (Les données régionales et nationales du Relevé des oiseaux nicheurs permettent-elles de bien prédire la tendance des populations de passereaux dans un secteur boréal vierge?) Avian Conservation and Ecology 9(1):5 (comprend un résumé en français).
- Machtans, C.S., C.H. Wedeles et E.M. Bayne. 2013. A first estimate for Canada of the number of birds killed by colliding with building windows (Première estimation canadienne du nombre d'oiseaux morts par collision avec les fenêtres de bâtiments). Avian Conservation and Ecology 8(2):6 (comprend un résumé en français).
- Mahony, N., E. Nol et T. Hutchinson. 1997. Food-chain chemistry, reproductive success, and foraging behaviour of songbirds in acidified maple forests of central Ontario. Revue canadienne de zoologie 75(4):509 517 (comprend un résumé en français).
- Main, A.R., J.V. Headley, K.M. Peru, N.L. Michel, A.J. Cessna et C.A. Morrissey. 2014. Widespread use and frequent detection of neonicotinoid insecticides in wetlands of Canada's prairie pothole region. PLoS One 9(3):e92821.
- Mänd, R., V. Tilgar et A. Leivits. 2000. Calcium, snails, and birds: a case study. Web Ecology 1:63 69.
- Manning Cooper and Associates Ltd. 2007. Canada Warbler (Wilsonia canadensis) Habitat Sampling in the Fort Nelson Forest. District Canadian Forest Products Ltd., Fort Nelson, BC.
- Masek, J.G., W.B. Cohen, D. Leckie, M.A. Wulder, R. Vargas, B. de Jong, S. Healey, B. Law, R. Birdsey et R. Houghton. 2011. Recent rates of forest harvest and conversion in North America. Journal of Geophysical Research: Biogeosciences (2005 2012) 116(G4).
- McClure, C. J., B.W. Rolek, K. McDonald et G.E. Hill. 2012. Climate change and the decline of a once common bird. Ecology and Evolution 2(2):370 378.
- McDermott, M.E., et A.D. Rodewald. 2014. Conservation value of silvopastures to Neotropical migrants in Andean forest flocks. Biological Conservation 175:140 147.
- McDonald, M.V. 2013. Kentucky Warbler (Geothlypis formosa). Cornell Lab of Ornithology, Ithaca, NY. (consulté en août 2014).
- McShea, W.J., M.V. McDonald, E.S. Morton, R. Meier et J.H. Rappole. 1995. Long-term trends in habitat selection by Kentucky Warblers. The Auk 112(2):375 381.
- Miller, N.A. 1999. Landscape and habitat predictors of Canada Warbler (Wilsonia canadensis) and Northern Waterthrush (Seiurus noveboracensis) occurrence in Rhode Island swamps. Thèse de maîtrise, University of Rhode Island Kingston, RI, États Unis.
- Mineau, P. 2005. Direct losses of birds to pesticides – beginnings of a quantification, p. 1065 1070 in Bird Conservation Implementation and Integration in the Americas: Proceedings of the Third International Partners in Flight Conference 2002. USDA Forest Service, Albany, CA, GTR-PSW-191.
- Mineau, P., et C. Palmer. 2013. The impact of the nation's most widely used insecticides on birds: neonicotinoid insecticides and birds. American Bird Conservancy, Washington D.C.
- Mineau, P., et M. Whiteside. 2013. Pesticide acute toxicity is a better correlate of US grassland bird declines than agricultural intensification. PLoS One 8(2):e57457.
- Møller, A.P., D. Rubolini et E. Lehikoinen. 2008. Populations of migratory bird species that did not show a phenological response to climate change are declining, Proceedings of the National Academy of Sciences 105(42):16195 16200.
- NatureServe. 2013. NatureServe Explorer: an online encyclopedia of life, version 7.1, NatureServe, Arlington, VA.
- Nebel, S., A. Mills, J.D. McCracken et P.D. Taylor. 2010. Declines of aerial insectivores in North America follow a geographic gradient (Présence d'un gradient géographique dans le déclin des insectivores aériens). Avian Conservation and Ecology 5(2):1 (comprend un résumé en français).
- Newson, S., S. Mendes, H. Crick, N. Dulvy, J. Houghton, G. Hays, A. Hutson, C. MacLeod, G. Pierce et R. Robinson. 2009. Indicators of the impact of climate change on migratory species. Endangered Species Research 7(2):101 113.
- Nocera, J.J., J.M. Blais, D.V. Beresford, L.K. Finity, C. Grooms, L.E. Kimpe, K. Kyser, N. Michelutti, M.W. Reudink et J.P. Smol. 2012. Historical pesticide applications coincided with an altered diet of aerially foraging insectivorous Chimney Swifts. Proceedings of the Royal Society B: Biological Sciences 279(1740):3114 3120.
- North American Bird Conservation Initiative US Committee. 2010. The state of the birds: 2010, report on climate change. United States of America, US Department of the Interior, Washington D.C.
- Norton, M., J. Bendell, L. Bendell-Young et C. LeBlanc. 2001. Secondary effects of the pesticide Bacillus thuringiensis kurstaki on chicks of Spruce Grouse (Dendragapus canadensis). Archives of environmental contamination and toxicology 41(3):369 373.
- Paquette, S.R., F. Pelletier, D. Garant et M. Bélisle. 2014. Severe recent decrease of adult body mass in a declining insectivorous bird population. Proceedings of the Royal Society B: Biological Sciences 281(1786):1 9.
- Partners in Flight Science Committee. 2012. Species Assessment Database. Version 2012, Laurel, MD. (consulté le 16 septembre 2014).
- Partners in Flight Science Committee. 2013. Population Estimates Database. Version 2013, Laurel, MD. (consulté le 16 septembre 2014).
- Portillo-Quintero, C., A. Sanchez, C. Valbuena, Y. Gonzalez et J. Larreal. 2012. Forest cover and deforestation patterns in the Northern Andes (Lake Maracaibo Basin): a synoptic assessment using MODIS and Landsat imagery. Applied Geography 35(1):152 163.
- Porvari, P., M. Verta, J. Munthe et M. Haapanen. 2003. Forestry practices increase mercury and methyl mercury output from boreal forest catchments, Environmental Science & Technology 37(11):2389 2393.
- Price, P.W., R.F. Denno, M.D. Eubanks, D.L. Finke et I. Kaplan. 2011. Insect ecology: behavior, populations and communities, Cambridge University Press, New York, NY, 812 p.
- Rappole, J.H., D.I. King et J.H.V. Rivera. 2003. Coffee and conservation. Conservation Biology 17(1):334 336.
- Reitsma, L., M. Goodnow, M.T. Hallworth et C.J. Conway. 2010. Canada Warbler (Cardellina canadensis), Cornell Lab of Ornithology, Ithaca, NY. (consulté le 17 septembre 2014).
- Rich, T.D., C.J. Beardmore, H. Berlanga, P.J. Blancher, M.S. Bradstreet, G.S. Butcher, D. Demarest, E.H. Dunn, W.C. Hunter, E.E. Iñigo Elias, J.A. Kennedy, A.M. Martell, A.O. Panjabi, D.N. Pashley, K.V. Rosenberg, C.M. Rustay, J.S. Wendt et T.C. Will. 2004. Partners in Flight North American Landbird Conservation Plan. Cornell Lab of Ornithology, Ithaca, NY.
- Rimmer, C.C., E.K. Miller, K.P. McFarland, R.J. Taylor et S.D. Faccio. 2010. Mercury bioaccumulation and trophic transfer in the terrestrial food web of a montane forest. Ecotoxicology 19(4):697 709.
- Rioux, S., J. P.L. Savard et A.A. Gerick. 2013. Avian mortalities due to transmission line collisions: a review of current estimates and field methods with an emphasis on applications to the Canadian electric network (Mortalité aviaire attribuable aux collisions avec les lignes de transport d'électricité : une revue des estimations actuelles et des méthodes de terrain avec un accent sur les applications au réseau électrique canadien). Avian Conservation and Ecology 8(2):7 (comprend un résumé en français).
- Robinson, A., H.Q. Crick, J.A. Learmonth, I.M. Maclean, C.D. Thomas, F. Bairlein, M.C. Forchhammer, C.M. Francis, J.A. Gill, B.J. Godley, J. Harwood, G.C. Hays, B. Huntley, A.M. Hutson, G.J. Pierce, M.M. Rehfisch, D.W. Sims, M.B. Santos, T.H. Sparks, D.A. Stroud et M.E. Visser. 2009. Travelling through a warming world: climate change and migratory species. Endangered Species Research 7:87 89.
- Robinson, S., J. Fitzpatrick et J. Terborgh. 1995. Distribution and habitat use of Neotropical migrant landbirds in the Amazon basin and Andes. Bird Conservation International 5(2-3):305 323.
- Roca, R., L. Adkins, M.C. Wurschy et K. Skerl. 1996. Transboundary conservation: an ecoregional approach to protect neotropical migratory birds in South America. Environmental Management 20(6):849 863.
- Rodenhouse, N., S. Matthews, K. McFarland, J. Lambert, L. Iverson, A. Prasad, T. Sillett et R. Holmes. 2008. Potential effects of climate change on birds of the Northeast. Mitigation and Adaptation Strategies for Global Change 13(5 6):517 540.
- Rooney, T.P. 2009. High white-tailed deer densities benefit graminoids and contribute to biotic homogenization of forest ground-layer vegetation. Plant Ecology 202(1):103 111.
- Russell, F.L., D.B. Zippin et N.L. Fowler. 2001. Effects of White-tailed deer (Odocoileus virginianus) on plants, plant populations and communities: a review. The American Midland Naturalist 146(1):1 26.
- Ryan, C.M. J. 2012. Mapping potential habitat for Canada Warblers (Cardellina canadensis) in Dartmouth, NS, mémoire de spécialisation de baccalauréat, Dalhousie University, Halifax, NS.
- Sager, T.A. 1997. Organochlorine pesticide contamination in New World passerines. Honors Project, Paper 10, Illinois Wesleyan University.
- Saino, N., R. Ambrosini, D. Rubolini, J. von Hardenberg, A. Provenzale, K. Hüppop, O. Hüppop, A. Lehikoinen, E. Lehikoinen et K. Rainio. 2011. Climate warming, ecological mismatch at arrival and population decline in migratory birds. Proceedings of the Royal Society B: Biological Sciences 278(1707):835 842.
- Sánchez-Cuervo, A.M., T.M. Aide, M.L. Clark et A. Etter. 2012. Land cover change in Colombia: surprising forest recovery trends between 2001 and 2010. PLoS One 7(8):e43943.
- Sanders, C.E., et D.J. Mennill. 2014a. Acoustic monitoring of migratory birds over western Lake Erie: avian responses to barriers and the importance of islands. The Canadian Field-Naturalist 128(2): 135 144.
- Sanders, C.E., et D.J. Mennill. 2014b. Acoustic monitoring of nocturnally migrating birds accurately assesses the timing and magnitude of migration through the Great Lakes. The Condor 116(3):371 383.
- Scheuhammer, A.M., M.W. Meyer, M.B. Sandheinrich et M.W. Murray. 2007. Effects of environmental methylmercury on the health of wild birds, mammals, and fish. AMBIO: A Journal of the Human Environment 36(1):12 19.
- Schieck, J., M. Nietfeid et J.B. Stelfox. 1995. Differences in bird species richness and abundance among three successional stages of aspen-dominated boreal forests. Revue canadienne de zoologie 73(8):1417 1431 (comprend un résumé en français).
- Schieck, J., et S.J. Song. 2006. Changes in bird communities throughout succession following fire and harvest in boreal forests of western North America: literature review and meta-analyses. Revue canadienne de recherche forestière 36(5):1299 1318 (comprend un résumé en français).
- Schieck, J., K. Stuart-Smith et M. Norton. 2000. Bird communities are affected by amount and dispersion of vegetation retained in mixedwood boreal forest harvest areas. Forest Ecology and Management 126(2):239 254.
- Schneider, R.R., J.B. Stelfox, S. Boutin et S. Wasel. 2003. Managing the cumulative impacts of land-uses in the Western Canadian Sedimentary Basin: a modeling approach. Conservation Ecology 7(1):8.
- Secretariat of the Stockholm Convention. 2011. United Nations targets widely-used pesticide endosulfan for phase out. (consulté en août 2014).
- Semenchuk, G. 2007. The Atlas of Breeding Birds of Alberta: A Second Look. The Federation of Alberta Naturalists, Edmonton, AB.
- Shannon, J.D. 1999. Regional trends in wet deposition of sulfate in the United States and SO2 emissions from 1980 through 1995. Atmospheric Environment 33(5):807 816.
- Sodhi, N.S., et C.A. Paszkowski. 1995. Habitat use and foraging behavior of four parulid warblers in a second-growth forest. Journal of Field Ornithology 66(2):277 288.
- Sopuck, L., K. Ovaska et B. Whittington. 2002. Responses of songbirds to aerial spraying of the microbial insecticide Bacillus thuringiensis var. kurstaki(Foray 48B®) on Vancouver Island, British Columbia, Canada. Environmental Toxicology and Chemistry 21(8):1664 1672.
- Stiles, F.G. 1994. A study of fall migration of Nearctic-breeding landbirds in central Costa Rica. Bird Conservation International 4(2-3):71 89.
- Stothart, P. 2011. F&F 2011: Fact$ and Figure$ of the Canadian Mining Industry. The Mining Association of Canada.
- Strode, P.K. 2003. Implications of climate change for North American wood warblers (Parulidae). Global Change Biology 9(8):1137 1144.
- Suarez, H.A., et G. Colorado. 2013. Densidad relativa de cuatro especies de aves migratorias y su relación con los atributos floisticos en diferentes usos del suelo en el municipio de Fredonia, Antioquia. Tesis de pregrado, Universidad de Antioquia (Université d'Antioquia), Medellín, Antioquia, Colombie.
- Thiffault, N., N.J. Fenton, A.D. Munson, F. Hébert, R.A. Fournier, O. Valeria, R.L. Bradley, Y. Bergeron, P. Grondin, et D. Paré. 2013. Managing understory vegetation for maintaining productivity in black spruce forests: a synthesis within a multi-scale research model. Forests 4(3):613 631.
- Titterington, R., H. Crawford et B. Burgason. 1979. Songbird responses to commercial clear-cutting in Maine spruce-fir forests. The Journal of Wildlife Management :602 609.
- U.S. Bureau of Land Management. 1978. Grass Creek: oil and gas leasing environmental assessment record. U.S. Bureau of Land Management, Worland District, WY.
- U.S. Environmental Protection Agency. 2010. Endosulfan phase-out. U.S. Environmental Protection Agency, [consulté en août 2014].
- U.S. Environmental Protection Agency. 2014. Environmental effects of acid rain, [consulté en août 2014].
- U.S. Geological Survey. 2000. Mercury in the environment, fact sheet 146-00, [consulté en août 2014].
- Van Wilgenburg, S.L., K.A. Hobson, E.M. Bayne et N. Koper. 2013. Estimated avian nest loss associated with oil and gas exploration and extraction in the Western Canadian Sedimentary Basin (Estimation de la perte de nids d'oiseaux attribuable à l'exploration et à l'exploitation pétrolière et gazière dans le bassin sédimentaire de l'Ouest canadien). Avian Conservation and Ecology 8(2):9 (comprend un résumé en français).
- Visser, M.E., L.J. Holleman et P. Gienapp. 2006. Shifts in caterpillar biomass phenology due to climate change and its impact on the breeding biology of an insectivorous bird. Oecologia 147(1):164 172.
- Vuori, K.-M., O. Siren et H. Luotonen. 2003. Metal contamination of streams in relation to catchment silvicultural practices: a comparative study in Finnish and Russian headwaters, Boreal environment research 8(1):61 70.
- Welsh, D.A., et D.R. Fillman. 1980. The impact of forest cutting on boreal bird populations. American Birds 34:84 94.
- Wiener, J.G., D.P. Krabbenhoft, G.H. Heinz et A.M. Scheuhammer. 2003. Ecotoxicology of mercury, p. 409 463, et al.,in D.J. Hoffman, B.A. Rattner, G.A. Burton, Jr. et J. Cairns, Jr. (éd.), Handbook of Ecotoxicology. 2e édition, Boca Raton, FL.
- Wolfe, J.D., M.D. Johnson et C.J. Ralph. 2014. Do birds select habitat or food resources? Nearctic-neotropic migrants in northeastern Costa Rica, PLoS One 9(1):e86221.
- Xing, Z., L. Chow, H. Rees, F. Meng, S. Li, B. Ernst, G. Benoy, T. Zha et L.M. Hewitt. 2013. Influences of sampling methodologies on pesticide-residue detection in stream water. Archives of environmental contamination and toxicology 64(2):208 218.
- Young, J.E., G.A. Sánchez-Azofeifa, S.J. Hannon et R. Chapman. 2006. Trends in land cover change and isolation of protected areas at the interface of the southern boreal mixedwood and aspen parkland in Alberta, Canada. Forest Ecology and Management 230(1 3):151 161.
- Zang, H. 1998. Auswirkungen des "Sauren Regens" (Waldsterben) auf eine Kohlmeisen-(Parus major-) Population in den Hochlagen des Harzes. Journal für Ornithologie 139(3):263 268
- Zimmerling, J.R., A.C. Pomeroy, M.V. d'Entremont et C.M. Francis. 2013. Canadian estimate of bird mortality due to collisions and direct habitat loss associated with wind turbine developments (Estimation de la mortalité aviaire canadienne attribuable aux collisions et aux pertes directes d'habitat associées à l'éolien). Avian Conservation and Ecology 8(2):10 (comprend un résumé en français).
- Zlonis, E.J., et G.J. Niemi. 2014. Avian communities of managed and wilderness hemiboreal forests. Forest Ecology and Management 328:26 34.
Communications personnelles
B. Drolet. 2014. biologiste, Environnement Canada, Québec, QC.
K. Fraser. 2014. Professeur adjoint, Université du Manitoba, Winnipeg, MB.
S. Haché. 2014. Biologiste des oiseaux terrestres, Environnement Canada, Yellowknife, NT.
A. Smith. 2014. Biostatisticien principal, Centre national de la recherche faunique, Ottawa, ON.
A. Westwood. 2014. Doctorant, Département de biologie, Dalhousie University, Halifax, NS.
S. Wilson. 2014. Direction générale des sciences et de la technologie, Environnement Canada, Ottawa, ON.
Annexe A : Effets sur l'environnement et sur les espèces non ciblées
Une évaluation environnementale stratégique (EES) est effectuée pour tous les documents de planification du rétablissement produits aux termes de la Loi sur les espèces en péril, conformément à La directive du Cabinet sur l'évaluation environnementale des projets de politiques, de plans et de programmes. Le but de cette évaluation est d'intégrer les facteurs environnementaux dans l'élaboration des politiques, plans et programmes publics, de manière à permettre une prise de décision éclairée, et de jauger si les résultats d'un document de planification du rétablissement pourront toucher tout élément de l'environnement ou influer sur les objectifs de la Stratégie fédérale de développement durable (SFDD).
La planification du rétablissement vise à favoriser les espèces en péril et la biodiversité en général. On reconnaît toutefois que, outre les avantages attendus, certains programmes peuvent avoir des conséquences imprévues sur l'environnement. Le processus de planification fondé sur des lignes directrices nationales tient compte directement de tous les effets sur l'environnement, en particulier des incidences possibles sur des espèces ou des milieux non ciblés. Les résultats de l'EES sont pris en compte dans le programme de rétablissement lui-même, et sont en outre résumés ci-dessous.
La Paruline du Canada niche souvent dans des milieux humides dotés d'une riche végétation dans un paysage boisé, de sorte que la protection de son habitat de reproduction peut passer par la protection des milieux humides (selon les préférences locales de l'espèce en matière d'habitat) et des vieilles forêts, ce qui profitera à d'autres espèces forestières. Par exemple, on observe la Paruline du Canada à des endroits où peuvent aussi être signalés la Paruline tigrée (Setophaga tigrina), la Chouette rayée (Strix varia) et le Moucherolle tchébec (Empidonax minimus). De même, les mesures visant à améliorer l'habitat d'hivernage peuvent entraîner des effets additifs qui favoriseront plusieurs autres espèces d'oiseaux terrestres en péril qui ont le même habitat d'hivernage que la Paruline du Canada ou dont l'aire d'hivernage chevauche celle de la Paruline du Canada, comme le Moucherolle à côtés olive (Contopus cooperi), le Pioui de l'Est (Contopus virens) (évalué par le COSEPAC mais ne figurant pas encore dans les listes de la LEP), la Paruline azurée (Setophaga caerulea), l'Engoulevent d'Amérique (Chordeiles minor), le Martinet ramoneur (Chaetura pelagica), le Moucherolle vert (Empidonax virescens), la Paruline à ailes dorées (Vermivora chrysoptera) et la Paruline orangée (Protonotaria citrea) (Robinson et al., 1995; Roca et al., 1996). De plus, les voies migratoires de la Paruline du Canada recoupent celles d'autres espèces en péril, comme le Goglu des prés (Dolichonyx oryzivorus), la Paruline azurée (Sanders et Mennill, 2014a), l'Hirondelle de rivage (Riparia riparia) et l'Hirondelle rustique (Hirundo rustica) (Stiles, 1994).
Cela dit, certaines espèces, y compris d'autres espèces en péril, préfèrent des conditions forestières différentes. Par exemple, deux oiseaux terrestres se reproduisent dans des milieux forestiers qui diffèrent de ceux qu'exploite la Paruline du Canada : le Pioui de l'Est, désigné « espèce préoccupante » par le COSEPAC, qui niche dans des peuplements forestiers matures ou intermédiaires à sous bois peu développé (COSEPAC, 2012), et l'Engoulevent bois-pourri, espèce menacée, qui niche dans des forêts semi-ouvertes présentant un sous-étage clair (COSEPAC, 2009). Les mesures de rétablissement de la Paruline du Canada doivent être coordonnées aux meilleures pratiques visant les autres espèces sauvages, dont les autres oiseaux chanteurs.
La possibilité que le présent programme de rétablissement entraîne des effets négatifs imprévus sur l'environnement et sur d'autres espèces a été examinée. La plupart des mesures recommandées sont non intrusives, y compris les relevés et les activités de sensibilisation du public. Il est donc peu probable que le programme de rétablissement ait des effets négatifs importants.
Annexe B : Autres activités de recherches visant les menaces connues et présumées qui pèsent sur la Paruline du Canada, ses proies, ainsi que sur leur habitat
La liste suivante présente certaines des études nécessaires pour comprendre les menaces qui pèsent sur l'espèce, ses proies, ainsi que sur leur habitat.
Perte ou dégradation d'habitat
- Étudier la vitesse à laquelle surviennent divers types de perte d'habitat ainsi que leurs répercussions dans l'ensemble des aires autres que les aires de reproduction.
- Déterminer l'importance relative de l'habitat de reproduction et de l'habitat autre que l'habitat de reproduction dans les baisses d'effectifs.
- Établir le volume (et les caractéristiques) des activités d'exploitation forestière et de sylviculture qui peut être réalisé tout en préservant suffisamment d'habitat convenable à l'échelle du paysage pour les populations de Parulines du Canada.
- Déterminer les effets des modifications des régimes hydrologiques sur la Paruline du Canada.
- Évaluer l'impact du pâturage exercé par bétail sur l'habitat de la Paruline du Canada à la limite sud de la zone boréale de l'Ouest canadien.
Modifications des systèmes naturels
- Déterminer les liens possibles entre la disponibilité d'insectes et les taux de reproduction des Parulines du Canada.
- Établir s'il existe un manque de synchronisme entre la période de reproduction et la période d'abondance maximale des proies.
- Déterminer les effets de la perte d'habitat (en particulier dans les aires d'hivernage) sur la disponibilité de proies pour la Paruline du Canada.
- Évaluer l'exposition aux pesticides, au mercure et à l'acidification des milieux dans l'ensemble de l'aire de répartition de la Paruline du Canada, ainsi que les effets possibles de cette exposition sur la disponibilité de proies.
Mortalité accidentelle
- Effectuer le suivi de la fréquence des collisions et déterminer les caractéristiques des sites associés à des taux de collision élevés.
Espèces envahissantes ou problématiques
- Étudier l'impact du parasitisme des couvées sur la productivité de la Paruline du Canada.
- Établir les risques de prédation associés à l'activité humaine dans les zones urbaines et rurales (p. ex. prédation par les chats et d'autres espèces dont les effectifs augmentent en raison de la présence humaine).
Changements climatiques et phénomènes météorologiques violents
- Déterminer les impacts des changements climatiques sur l'espèce et son habitat.
Pollution
- Évaluer l'exposition de la Paruline du Canada à la pollution (pesticides et mercure) et en préciser les impacts.
- Déterminer si l'acidification des milieux que fréquente l'espèce a un effet négatif sur la Paruline du Canada et son habitat (p. ex. en raison de la perte de sites de nidification ou d'alimentation de prédilection, de l'augmentation du risque de prédation et d'une carence en calcium durant les périodes de ponte et d'élevage des oisillons).
Détails de la page
- Date de modification :