
Bécasseau maubèche (Calidris canutus) : programme de rétablissement et plan de gestion 2017
Loi sur les espèces en péril
Série de programmes de rétablissement
Programme de rétablissement
Calidris canutus rufa
Calidris canutus roselaari
Plan de gestion
Calidris canutus islandica

Photo : © Jan van de Kam
2017
Table des matières
- Table des matières
- Préface
- Remerciements
- Sommaire
- Résumé du caractère réalisable du rétablissement
- 1. Évaluation de l'espèce par le COSEPAC
- 2. Information sur la situation de l'espèce
- 3. Information sur l'espèce
- 4. Menaces
- 5. Objectifs en matière de population et de répartition (rufa et roselaari)/objectifs de gestion (islandica)
- 6. Stratégies et approches générales pour l'atteinte des objectifs
- 7. Habitat essentiel
- 8. Mesure des progrès
- 9. Énoncé sur les plans d'action
- 10. Références
- Annexe A: Effets sur l'environnement et sur les espèces non ciblées
- Annexe B: Besoins en recherche
Information sur le document
Programme de rétablissement et plan de gestion du Bécasseau maubèche (Calidris canutus) au Canada - 2017
Loi sur les espèces en péril
Série de programmes de rétablissement
Programme de rétablissement
Calidris canutus rufa
Calidris canutus roselaari
Plan de gestion
Calidris canutus islandica

Référence recommandée
Environnement et Changement climatique Canada. 2017. Programme de rétablissement et Plan de gestion du Bécasseau maubèche (Calidris canutus) au Canada, Série de Programmes de rétablissement de la Loi sur les espèces en péril, Environnement et Changement climatique Canada, Ottawa, ix + 71 p.
Pour télécharger le présent programme de rétablissement ou pour obtenir un complément d'information sur les espèces en péril, incluant les rapports de situation du Comité sur la situation des espèces en péril au Canada (COSEPAC), les descriptions de la résidence, les plans d'action et d'autres documents connexes portant sur le rétablissement, veuillez consulter le Registre public des espèces en péril.
Illustration de la couverture : Bécasseau maubèche © Jan van de Kam
Also available in English under the title “Recovery Strategy and Management Plan for the Red Knot (Calidris canutus) in Canada”
Le contenu du présent document (à l'exception des illustrations) peut être utilisé sans permission, mais en prenant soin d'indiquer la source.
Préface
En vertu de l'Accord pour la protection des espèces en péril (1996), les gouvernements fédéral, provinciaux et territoriaux signataires ont convenu d'établir une législation et des programmes complémentaires qui assureront la protection efficace des espèces en péril partout au Canada. En vertu de la Loi sur les espèces en péril (L.C. 2002, ch. 29) (LEP), les ministres fédéraux compétents sont responsables de l'élaboration des programmes de rétablissement pour les espèces inscrites comme étant disparues du pays, en voie de disparition ou menacées, et des plans de gestion pour les espèces inscrites comme étant préoccupantes. Ils sont également tenus de rendre compte des progrès réalisés dans les cinq ans suivant la publication du document final dans le Registre public des espèces en péril.
La ministre de l'Environnement et du Changement climatique et ministre responsable de l'Agence Parcs Canada est le ministre compétent en vertu de la LEP à l’égard du Bécasseau maubèche et a élaboré ce document, conformément aux articles 37 et 65 de la LEP. Dans la mesure du possible, le programme de rétablissement a été préparé en collaboration avec la Colombie-Britannique, l'Alberta, la Saskatchewan, le Manitoba, l'Ontario, le Québec, le Nouveau-Brunswick, l'Île-du-Prince-Édouard, la Nouvelle-Écosse et Terre-Neuve-et-Labrador, les territoires du Yukon, du Nunavut et des Territoires du Nord-Ouest et tout autre personne ou organisation définie en vertu des paragraphes 39(1) et 66(1) de la LEP.
La réussite du rétablissement et/ou de la conservation de l'espèce dépendra de l'engagement et de la collaboration d'un grand nombre de parties concernées qui participeront à la mise en œuvre des directives formulées dans le présent document. Cette réussite ne pourra reposer seulement sur Environnement et Changement climatique Canada et l'Agence Parcs Canada ou sur toute autre autorité responsable. Tous les Canadiens et les Canadiennes sont invités à appuyer le document et à contribuer à sa mise en œuvre pour le bien du Bécasseau maubèche et de l'ensemble de la société canadienne.
Le présent document sera suivi d'un ou de plusieurs plans d'action pour les sous-espèces rufa et roselaari du Bécasseau maubèche qui présenteront de l'information sur les mesures de rétablissement qui doivent être prises par Environnement et Changement climatique Canada et l'Agence Parcs Canada et d'autres autorités responsables et/ou organisations participant à la conservation de l'espèce. La mise en œuvre du présent document est assujettie aux crédits, aux priorités et aux contraintes budgétaires des autorités responsables, organisations et conseils de gestion des ressources fauniques participants.
Le programme de rétablissement établit l’orientation stratégique visant à arrêter ou à renverser le déclin de l’espèce, incluant la désignation de l’habitat essentiel dans la mesure du possible. Il fournit à la population canadienne de l’information pour aider à la prise de mesures visant la conservation de l’espèce. Lorsque l’habitat essentiel est désigné, dans un programme de rétablissement ou dans un plan d’action, la LEP exige que l’habitat essentiel soit alors protégé.
Dans le cas de l’habitat essentiel désigné pour les espèces terrestres, y compris les oiseaux migrateurs, la LEP exige que l’habitat essentiel désigné dans une zone protégée par le gouvernement fédéralNote1de bas de page soit décrit dans la Gazette du Canada dans un délai de 90 jours après l’ajout dans le Registre public du programme de rétablissement ou du plan d’action qui a désigné l’habitat essentiel. L’interdiction de détruire l’habitat essentiel aux termes du paragraphe 58(1) s’appliquera 90 jours après la publication de la description de l’habitat essentiel dans la Gazette du Canada.
Pour l’habitat essentiel se trouvant sur d’autres terres domaniales, le ministre compétent doit, soit faire une déclaration sur la protection légale existante, soit prendre un arrêté de manière à ce que les interdictions relatives à la destruction de l’habitat essentiel soient appliquées.
Si l’habitat essentiel d’un oiseau migrateur ne se trouve pas dans une zone protégée par le gouvernement fédéral, sur le territoire domanial, à l’intérieur de la zone économique exclusive ou sur le plateau continental du Canada, l’interdiction de le détruire ne peut s’appliquer qu’aux parties de cet habitat essentiel -- constituées de tout ou partie de l’habitat auquel la Loi de 1994 sur la convention concernant les oiseaux migrateurs s’applique aux termes des paragraphes 58(5.1) et 58(5.2) de la LEP.
En ce qui concerne tout élément de l’habitat essentiel se trouvant sur le territoire non domanial, si le ministre compétent estime qu’une partie de l’habitat essentiel n’est pas protégée par des dispositions ou des mesures en vertu de la LEP ou d’autre loi fédérale, ou par les lois provinciales ou territoriales, il doit, comme le prévoit la LEP, recommander au gouverneur en conseil de prendre un décret visant l’interdiction de détruire l’habitat essentiel. La décision de protéger l’habitat essentiel se trouvant sur le territoire non domanial et n’étant pas autrement protégé demeure à la discrétion du gouverneur en conseil.
Remerciements
Le présent document a été préparé par Julie McKnight (Environnement et Changement climatique Canada, Service canadien de la faune [ECCC SCF] – Région de l'Atlantique) avec la généreuse contribution de Garry Donaldson (ECCC SCF – Région de l'Atlantique) et des membres du Comité technique sur les oiseaux de rivage d'Environnement et Changement climatique Canada, particulièrement Cheri Gratto-Trevor (ECCC – Direction générale de la science et de la technologie (ECCC S T) – Recherche sur les espèces sauvages), Anne McKellar (ECCC-SC - Région des Prairies et du Nord), Jennie Rausch (ECCC SCF – Région des Prairies et du Nord), Christian Friis (ECCC SCF – Région de l'Ontario), R.I.G. Morrison (ECCC S T, Recherche sur les espèces sauvages), Cynthia Pekarik (ECCC-SCF - Région de la capitale nationale), Paul Smith (ECCC S T – Recherche sur les espèces sauvages), Yves Aubry (ECCC SCF – Région du Québec) et Julie Paquet (ECCC SCF – Région de l'Atlantique). Nous remercions Kristiina Ovaska d'avoir facilité l'appel avec des experts du calculateur de menaces pour le rufa, David Andrews (ECCC SCF – Région de l'Atlantique) d'avoir réalisé les cartes incluses dans le présent document et Patricia M. González pour avoir fourni des conseils sur une version antérieure. Nous offrons aussi notre reconnaissance et nos remerciements à toutes les autres parties qui ont fourni des conseils et des renseignements ayant contribué à l'élaboration du présent document, y compris divers organismes et particuliers autochtones, des gouvernements provinciaux et territoriaux, d'autres ministères fédéraux (p. ex. Agence Parcs Canada), des propriétaires fonciers, des citoyens et divers intervenants.
Sommaire
Le Bécasseau maubèche (Calidris canutus) est un oiseau de rivage de taille moyenne présentant un profil de bécasseau typique : un long bec et une tête assez petite, de longues ailes effilées donnant au corps un profil allongé et fuselé, et de longues pattes. Le plumage nuptial du Bécasseau maubèche est très caractéristique : la face, le cou, la poitrine et la majorité du dessous prenant une coloration roussâtre distinctive. Trois sous-espèces se rencontrent au Canada : Calidris canutus rufa (ci-après « rufa ») se reproduit uniquement au Canada, Calidris canutus islandica (ci-après « islandica ») se reproduit au Canada et au Groenland, et Calidris canutus roselaari (ci-après « roselaari ») se reproduit en Alaska et en Russie, et fréquente, en petits nombres, le territoire canadien pendant la migration. En raison de leur déclin à long terme, le rufa est inscrit comme espèce « en voie de disparition », le roselaari, comme espèce « menacée », et l'islandica, comme espèce « préoccupante » à l'annexe 1 de la LEP. Depuis l'évaluation de 2007 comme espèce « en voie de disparition » par le Comité sur la situation des espèces en péril au Canada (COSEPAC), de nouvelles données sur le type roselaari donnent à penser que ce dernier ne se reproduit pas au Canada (les roselaari réputés nicher au Canada étaient en fait des rufa), et seules quelques haltes migratoires de faible importance ont été repérées au Canada. La totalité de la population mondiale de rufa, estimée à 42 000 individus, niche au Canada. Moins de 1 % de la population mondiale actuelle de roselaari, estimée à 17 000 individus, est réputée fréquenter le Canada pendant la migration, et environ 18 % de la population mondiale d'islandica, estimée à 450 000 individus, niche au Canada.
Le Bécasseau maubèche niche sur le sol, dans la toundra sèche et légèrement élevée (généralement à une altitude inférieure à 150 m au-dessus du niveau de la mer), à moins de 500 m d'un milieu humide d'eau douce ou d'un plan d'eau douce (lac, ruisseau, rivière ou étang). Pendant les périodes de migration et l'hiver, l'espèce a besoin d'un habitat (généralement des milieux côtiers marins ou estuariens mais aussi des lacs salés intérieurs pour l'alimentation et le repos) relativement exempt de perturbations anthropiques. Elle utilise les plages sablonneuses, flèches littorales, bancs de sable, battures de vase, restingas (c'est-à-dire plateformes rocheuses intertidales érodées par les vagues), plaines rocheuses intertidales et marais salés des haltes migratoires (Niles et al., 2007). Les haltes migratoires doivent donner accès à de la nourriture abondante et facile à digérer. Pendant la migration printanière dans la baie du Delaware (Delaware et New Jersey, aux États-Unis), la présence de limules (Limulus polyphemus) en fraye est essentielle pour la sous-espèce rufa. En effet, les œufs de limules constituent une source de nourriture vitale. Le Bécasseau maubèche hiverne le long de plages sablonneuses, mais utilise également des berges rocheuses, des restingas, des plaines rocheuses intertidales, des bancs de tourbe, des marais salés, des rizières, des lagunes saumâtres et des battures de vase.
Les menaces qui pèsent sur l'espèce font partie des catégories du premier niveau de l'IUCN-CMP suivantes : développement résidentiel et commercial; agriculture et aquaculture; production d'énergie et exploitation minière; utilisation des ressources biologiques; intrusions et perturbations humaines; modifications des systèmes naturels (p. ex. gestion de l'eau et exploitation de barrages, stabilisation des berges); espèces et gènes envahissants ou autrement problématiques; pollution; changements climatiques et phénomènes météorologiques violents.
Le caractère réalisable du rétablissement du rufa et du roselaari comporte des inconnues. Conformément au principe de précaution, un programme de rétablissement a été élaboré, tel qu'il convient de le faire lorsque le rétablissement est déterminé comme étant réalisable. Malgré ces inconnues et suivant le principe de précaution, le présent document a été élaboré, conformément au paragraphe 41(1) de la LEP. Puisque le caractère réalisable du rétablissement ne s'applique pas aux espèces préoccupantes, il n'est pas établi pour l'islandica dans le présent document.
L'objectif à court terme en matière de population pour les sous-espèces rufa et islandica au Canada est de mettre fin au déclin de leur population d'ici 2025 à l'échelle du pays. L'objectif à long terme en matière de population pour le rufa sera ensuite d'augmenter, puis de maintenir la population à un niveau équivalent ou supérieur à ceux de 1986-1990 (100 000-150 000 individus). L’objectif à long terme en matière de population pour le rufa sera ensuite d’augmenter, puis de maintenir la population à un niveau équivalent ou supérieur à ceux de 1986-1990 (100 000-150 000 individus). L’objectif à long terme en matière de population pour l’islandica est de maintenir la population à son niveau actuel. Compte tenu des nouvelles données sur le roselaari depuis l’évaluation du COSEPAC, l’objectif est de conserver le roselaari au Canada et toutes ses haltes migratoires canadiennes fréquentées par au moins 1 % de la population actuelle (1 % = 170 individus), ce qui assurerait sa persistance en tant que migrateur au Canada.
Les stratégies générales à appliquer pour contrer les menaces à la survie et au rétablissement du Bécasseau maubèche sont présentées à la section 6.1 – Orientation stratégique pour le rétablissement.
Aux termes de la LEP, la désignation et la protection de l'habitat essentiel s'appliquent uniquement aux espèces en voie de disparition et aux espèces menacées. L'habitat essentiel à la survie et au rétablissement du rufa et du roselaari est partiellement désigné à la section 7.1. Puisque l'habitat essentiel ne s'applique pas aux espèces préoccupantes, il n'est pas désigné dans le cas de l'islandica dans le présent document. Un calendrier des études a été élaboré pour obtenir l'information nécessaire à la désignation complète de l'habitat essentiel en vue de l'atteinte des objectifs en matière de population et de répartition.
Un ou plusieurs plans d'action visant le rufa et le roselaari seront publiés dans le Registre public des espèces en péril dans les cinq ans suivant la publication du présent document.
Résumé du caractère réalisable du rétablissement
D'après les quatre critères suivants qu'Environnement et Changement climatique Canada utilise pour définir le caractère réalisable du rétablissement, le rétablissement du rufa et du roselaari comporte des inconnues. Conformément au principe de précaution, un programme de rétablissement a été élaboré en vertu du paragraphe 41(1) de la LEP, tel qu'il convient de faire lorsque le rétablissement est déterminé comme étant réalisable du point de vue technique et biologique. Le présent programme de rétablissement traite des inconnues entourant le caractère réalisable du rétablissement.
Puisque le caractère réalisable du rétablissement ne s'applique pas aux espèces préoccupantes, il n'est pas établi pour le C. c. islandica dans le présent document.
Des individus de l'espèce sauvage capables de se reproduire sont disponibles maintenant ou le seront dans un avenir prévisible pour maintenir la population ou augmenter son abondance.
C. c. rufa
Oui. La population du rufa était estimée à environ 42 000 individus en 2012 (Andres et al., 2012) et le rufa se rencontre actuellement dans l'ensemble de son aire de reproduction connue.C. c. roselaari
Oui. En 2012, la population était estimée à environ 17 000 individus (Andres et al., 2012; Carmona et al., 2013), qui nichent dans le nord-ouest et le nord de l'Alaska, aux États-Unis, et à l'île Wrangel, en Russie (Buchanan et al., 2010, 2011; Andres et al., 2012; Carmona et al., 2013). Selon les nouvelles données détaillées dans Andres et al. (2012) et Carmona et al. (2013), le roselaari ne nicherait pas au Canada, et seul un petit nombre d'individus (moins de 1 % de la population actuelle) utilisent les haltes migratoires en Colombie-Britannique (Carmona et al., 2013) pendant la migration vers le nord.De l'habitat convenable suffisant est disponible pour soutenir l'espèce, ou pourrait être rendu disponible par des activités de gestion ou de remise en état de l'habitat.
Le Bécasseau maubèche est possiblement l'une des espèces les plus difficiles à recenser en Arctique en raison de sa faible densité dans une région vaste et éloignée et de son comportement discret durant la nidification.
C. c. rufa
Oui. Il n'y a pas de preuve que l'habitat de reproduction convenable constitue un facteur limitatif pour l'espèce dans les grandes étendues de l'Arctique canadien. Des habitats de halte migratoire et d'hivernage convenables et suffisants pourraient être disponibles actuellement et davantage pourrait devenir disponible grâce à des mesures de gestion et/ou de restauration de l'habitat.C. c. roselaari
Oui. Il n'y a pas de preuve que l'habitat de reproduction convenable constitue un facteur limitatif pour l'espèce dans le nord-ouest et le nord de l'Alaska, aux États-Unis, et dans l'île Wrangel, en Russie. Le C. c. roselaari ne niche pas au Canada, et un nombre peu élevé d'individus utilisent les haltes migratoires canadiennes (les sites sont fréquentés par moins de 1 % de la population actuelle) (Carmona et al., 2013). La sous-espèce contourne généralement le Canada pendant la migration (U.S. Fish and Wildlife Service, 2011).Les principales menaces pesant sur l'espèce ou sur son habitat (y compris les menaces à l'extérieur du Canada) peuvent être évitées ou atténuées.
Le Bécasseau maubèche et d'autres oiseaux de rivage continuent d'être menacés par la chasse légale et illégale pratiquée dans les Caraïbes et des portions de l'Amérique du Sud. On ne sait pas vraiment si les populations de Bécasseaux maubèches se sont un jour rétablies de la chasse intensive des années 1800 qui a considérablement réduit leur abondance (Harrington, 2001; Cohen et al., 2009; Karpanty et al., 2014). Des activités visant à réglementer et/ou à interdire la chasse sont en cours dans certaines régions (p. ex. Barbade, Guadeloupe, Guyane française) et, selon les résultats attendus décrits dans l'évaluation des menaces de l'U.S. Fish and Wildlife Service (2014), la chasse au C .c. roselaari continuera à diminuer.
C. c. rufa
Inconnu. Une menace principale qui pèse sur la sous-espèce concerne la gestion de la pêche au limule le long de la côte de l’Atlantique des États-Unis. La surpêche des limules a privé les bécasseaux en migration d’une source de nourriture essentielle dont ils ont besoin pour récupérer après de longs vols, stocker des nutriments, augmenter leur masse corporelle en vue de la migration vers l'Arctique et faire des réserves supplémentaires favorisant leur survie après l'arrivée dans les lieux de reproduction (Morrison, 2006; Morrison et al., 2007). La restriction de la pêche de limules devrait permettre le rétablissement de leurs populations, ce qui pourrait, par le fait même, favoriser le rétablissement des effectifs de Bécasseaux maubèches puisqu'un lien a été établi entre la survie de ces derniers et la masse corporelle des individus au moment de leur départ de la baie du Delaware (Baker et al., 2004; McGowan et al., 2011).La dégradation des habitats ne servant pas à la reproduction et situés en dehors des limites du Canada ainsi que les perturbations qui y sévissent sont des menaces qui pourraient sans doute être atténuées, notamment grâce à l'intérêt pour la conservation et les projets/initiatives en cours à l'échelle internationale. Il est peut-être impossible d'atténuer les changements climatiques ainsi que les changements d'habitat qui s'ensuivent.
C. c. roselaari
Inconnu. La manière dont la sous-espèce utilise les haltes migratoires pendant sa migration automnale n'est pas bien comprise (U.S. Fish and Wildlife Service, 2011). La dégradation des habitats ne servant pas à la reproduction et situés en dehors des limites du Canada (tels que ceux de la baie de San Francisco et de Grays Harbor, dans l'État de Washington) ainsi que les perturbations qui y sévissent sont des menaces probables pour le roselaari (COSEWIC, 2007). On présume que ces menaces pourraient être atténuées.Des techniques de rétablissement existent pour atteindre les objectifs en matière de population et de répartition ou leur élaboration peut être prévue dans un délai raisonnable.
C. c. rufa
Inconnu. La gestion durable de la pêche au limule et la gestion des haltes migratoires importantes de manière à ce qu’elles subviennent aux besoins des oiseaux de rivage favoriseront le rétablissement continu. On ne sait pas très bien si les menaces potentielles extérieures au Canada, que des travaux de recherche pourraient confirmer, peuvent être évitées.C. c. roselaari
Inconnu. La petite population canadienne n’est présente que pendant la migration, et la vaste majorité de sa répartition et de sa population se trouve dans les lieux de reproduction (nord-ouest et nord de l’Alaska, aux États-Unis, et île Wrangel, en Russie) et d’hivernage (nord-ouest du Mexique). On ne sait pas très bien si les menaces potentielles extérieures au Canada, que des travaux de recherche pourraient confirmer, peuvent être évitées.
1. Évaluation de l'espèce par le COSEPACa
| Date de l'évaluation : | Nom commun (population) : | Nom scientifique : | Statut selon le COSEPAC : | Présence au Canada : | Historique du statut selon le COSEPAC : | Justification de la désignation |
|---|---|---|---|---|---|---|
| Avril 2007 | Bécasseau maubèche de la sous-espèce rufa | Calidris canutus rufa | En voie de disparition | NT, NU, BC, AB, SK, MB, ON, QC, NB, PE, NS, NL | Désignations attribuées en avril 2007 | Cette sous-espèce est un oiseau de rivage de taille moyenne, qui se reproduit seulement dans l'Arctique canadien et qui migre sur des milliers de kilomètres entre ses aires arctiques de reproduction et ses aires d'hivernage à la pointe de l'Amérique du Sud. Cette sous-espèce a subi un déclin de 70 % de son abondance au cours des trois dernières générations (15 ans). Elle est menacée par la pénurie d'œufs de limules, aliment essentiel durant la migration vers le nord. Aucune immigration à partir d'autres populations n'est possible. |
| Avril 2007 | Bécasseau maubèche du type roselaarib | Calidris canutus roselaari type | Menacée | YT, NT, BC | Désignations attribuées en avril 2007 | Cette unité désignable comprend la sous-espèce roselaari ainsi que deux autres populations qui hivernent en Floride et dans le nord du Brésil et qui semblent posséder des caractéristiques de roselaari. La sous-espèce roselaari passe par la Colombie-Britannique au cours de sa migration et elle se reproduit en Alaska. Les voies de migration et les aires de reproduction des deux autres populations sont inconnues. Ce groupe a subi un déclin global de 47 % au cours des trois dernières générations (15 ans). Parmi les menaces courantes, on compte la perte et la dégradation de l'habitat dans les aires d'hivernage et, dans le cas des groupes de la Floride et du sud-est des États-Unis et de Maranhão, une pénurie d'oeufs de limules, aliment essentiel durant la migration vers le nord. Une immigration à partir d'autres populations n'est pas prévue. |
| Avril 2007 | Bécasseau maubèche de la sous-espèce islandica | Calidris canutus islandica | Préoccupante | NT, NU | Désignations attribuées en avril 2007 | Cette sous-espèce est un oiseau de rivage de taille moyenne, qui se reproduit dans l'Arctique et qui migre vers ses aires d'hivernage situées en Europe. Quarante pour cent de la population reproductrice de cette sous-espèce se trouve au Canada. Elle a subi un déclin de 17 % au cours des trois dernières générations (15 ans). Les individus au Canada ne semblent être confrontés à aucune menace. Bien que son habitat dans les aires de reproduction au Canada soit probablement stable, la récolte de mollusques dans les aires d'hivernage en Europe représente une menace permanente. |
a COSEPAC (Comité sur la situation des espèces en péril au Canada)
b Voir la section 2 pour un résumé des nouvelles données sur cette sous espèce depuis la dernière évaluation du COSEPAC.
2. Information sur la situation de l'espèce
Dans le présent document, les termes « hiver », « hiverner » et « hivernage » sont utilisés pour désigner la période en dehors de la période de reproduction (aussi tôt que septembre et aussi tard que mai, mais généralement de décembre à février), soit quand les oiseaux ne sont pas en migration (selon le U.S. Fish and Wildlife Service, 2014).
De nouvelles données sur la répartition et l’abondance de la population de Calidris canutus roselaari (ci-après « roselaari ») ont été obtenues depuis l’évaluation du Bécasseau maubèche (Calidris canutus) par le COSEPAC, en 2007. Selon les résultats d’activités de baguage et de géolocalisation ainsi que des travaux antérieurs sur les isotopes stables (Atkinson et al., 2005), les Bécasseaux maubèches non nicheurs, que l’on prenait autrefois pour des roselaari le long de la côte ouest de la Floride, du sud-est des États-Unis et du nord du Brésil, sont probablement des Calidris canutus rufa (ci-après « rufa ») (Niles et al., 2008; Andres et al., 2012), et la quasi-totalité, voire la totalité, des Bécasseaux maubèches non nicheurs dans le nord-ouest du golfe du Mexique sont aussi des rufa (U.S. Fish and Wildlife Service, 2014). Ces données récentes indiquent que les roselaari sont principalement confinés à la côte du Pacifique de l’Amérique du Nord et de l’Amérique du Sud. Cette sous-espèce ne se reproduit pas dans l’ouest de l’Arctique canadien, contrairement à ce que l’on croyait et des liens évidents ont été tracés entre différents sites d’hivernage du nord-ouest du Mexique, des haltes migratoires dans l’État de Washington aux États-Unis et des aires de reproduction dans le nord-ouest et le nord de l’Alaska ainsi que sur l’île Wrangel en Russie (Buchanan et al., 2010, 2011; U.S. Fish and Wildlife Service, 2011; Andres et al., 2012; Carmona et al., 2013). Sa présence est considérée comme accidentelle au Yukon (Environment Yukon, 2014). Le U.S. Fish and Wildlife Service (2011) indique que la sous-espèce roselaari contourne le plus souvent la Colombie-Britannique pendant la migration.
On sait que l’ensemble de la population mondiale de rufa, estimée à 42 000 individus, se reproduit au Canada. On estime que moins de 1 % de la population mondiale actuelle de roselaari, estimée à 17 000 individus, fréquente le Canada durant sa migration et qu’environ 18 % de la population mondiale de Calidris canutus islandica (ci-après islandica), estimée à 450 000 individus, se reproduit au Canada (Wetlands International, 2015.)
Le rufa est inscrit comme espèce « en voie de disparition », le roselaari comme espèce « menacée » et l’islandica comme espèce « préoccupante » à l’annexe 1 de la Loi sur les espèces en périf (LEP) du gouvernement fédéral. Le tableau 1 présente les cotes de conservation du Bécasseau maubèche. L’Ontario, le Nouveau-Brunswick, la Nouvelle-Écosse et Terre-Neuve-et-Labrador ont inscrit le rufa dans leurs lois sur les espèces en péril respectives. Au Québec, le rufa est inscrit à la Liste des espèces de la faune vertébrée susceptibles d’être désignées menacées ou vulnérables. Cette liste est produite aux termes de la Loi sur les espèces menacées ou vulnérables (RLRQ, chap. E-12.01). L’islandica et le roselaari ne sont inscrits dans aucune loi provinciale ou territoriale relative aux espèces en péril.
Aux États-Unis, le rufa a été inscrit parmi les espèces menacées (« Threatened ») aux termes de l’Endangered Species Act des États-Unis en 2014. À l’échelle des États, la sous-espèce est menacée (« Threatened ») au New Jersey et préoccupante (« Special concern ») en Géorgie (Niles et al., 2005). En 2005, le rufa a été ajouté à l’annexe 1 de la Convention sur la conservation des espèces migratrices (CMS, ou Convention de Bonn; CMS, 2005), sur laquelle figurent les espèces migratrices menacées de disparition. Le Bécasseau maubèche a été inscrit comme espèce en danger critique (Critically Endangered) sur la liste rouge du ministère de l’Environnement du Brésil en 2014 et a été désigné espèce en danger (Endangered) en Argentine (López-Lanús et al., 2008, Resolución 348/2010, Secretaría de Ambiente y Desarrollo Sustentable). Au Chili, il a reçu cette même désignation (en danger) du Ministerio de la Secretaría General de la Presidencia de Chile en 2008. En Uruguay, la Dirección Nacional de Medio Ambiente considère également l’espèce comme en danger (Azpiroz et al., 2012) et prioritaire du point de vue de la conservation (Aldabe et al., 2013). Le Bécasseau maubèche figure sur la liste rouge en tant qu’espèce gravement en péril du ministère de l’Environnement du Brésil depuis 2014. L’Union internationale pour la conservation de la nature (IUCN) désigne le Bécasseau maubèche comme espèce de préoccupation mineure, mais ne précise pas le statut des six sous-espèces, lequel pourrait différer d’une sous-espèce à l’autre (BirdLife International, 2012).
| Sous-espèce | Cote mondiale (G)c | Cote nationale (N)d | Cote infranationale (S)e |
|---|---|---|---|
| rufa | G4T2 | Canada : N1B, N3N4N, N3M États-Unis : N1B |
Territoires du Nord-Ouest (S1B) Nunavut (SNRB) Colombie-Britannique (SNR) Alberta (SU) Saskatchewan (S2M) Manitoba (SNA) Ontario (S1N) Québec (S1M) Nouveau-Brunswick (S2M) Île-du-Prince-Édouard (S2M) Nouvelle-Écosse (S2S3M) Terre-Neuve (S3N) et Labrador (S3N) |
| roselaari | G4TNR | Canada : NNR | Yukon (SNA) Territoires du Nord-Ouest (SNR) Colombie-Britannique (SNR) |
| Islandica | G4TNR | Canada : N3B | Territoires du Nord-Ouest (S2B) Nunavut (SNRB) |
c Cote G -- Cote de conservation mondiale : G4 = espèce apparemment non en péril; T2 = sous-espèce en péril; TNR = sous-espèce non classée.
d Cote N -- Cote de conservation nationale : N1 = population au Canada est gravement en péril; N3 = population au Canada est vulnérable; N4 = population au Canada est apparemment non en péril; NNR = espèce non classée. B = population reproductrice; N = population non reproductrice; M = population migratrice.
e Cote S --Cote infranationale (provinciale ou territoriale) : S1 = espèce gravement en péril; S2 = espèce en péril; S3 = population vulnérable; S4 = espèce apparemment non en péril; S5 = espèce non en péril; U = espèce non classable; NR = espèce non classée; NA = non applicable. B = population reproductrice; N = population non reproductrice; M = population migratrice.
3. Information sur l'espèce
3.1 Description de l'espèce
Le Bécasseau maubèche est un oiseau de rivage de taille moyenne qui présente les caractéristiques morphologiques typiques du genre Calidris : bec long et petite tête, ailes longues et effilées donnant au corps un profil allongé et fuselé, pattes longues. Le plumage nuptial du Bécasseau maubèche est très caractéristique : la face, le cou, la poitrine et la majorité du dessous prenant une coloration roussâtre distinctive. Les plumes du dessus sont brun foncé ou noires, avec du roux et du gris, ce qui donne au dos une apparence pailletée. En plumage d'hiver, le Bécasseau maubèche est plus terne, sa face ventrale étant blanche, et son dos, gris pâle. Les six sous-espèces sont actuellement reconnues dans le monde. Les sous-espèces sont semblables en apparence, mais la taille du corps, la longueur du bec et le plumage varient légèrement (Baker et al., 2013); les sous-espèces forment des populations biogéographiques qui se distinguent quant à leur répartition et cycle annuel. Les sous-espèces qui se reproduisent au Canada sont le rufa et l'islandica. Au Canada, le roselaari se rencontre en petits nombres pendant la migration.
3.2 Population et répartition de l'espèce

Description longue pour la figure 1
La figure 1 montre, à l’échelle mondiale, les aires de reproduction et de migration ainsi que les quartiers d’hiver/aires utilisées en dehors de la période de reproduction, les voies migratoires et les sites importants utilisés en dehors de la période de reproduction des Bécasseaux maubèches des sous espèces rufa, roselaari et islandica. Le rufa se reproduit dans l’île de Baffin et dans les îles situées à l’ouest de cette dernière et au sud des îles Devon, Cornwallis, Bathurst, Byam Martin et Melville. Le rufa migre à travers l’est et le centre du Canada et le long de la côte est des États Unis, vers le sud jusque dans la moitié sud de cette dernière et sur les côtes du golfe du Mexique, de l’Amérique centrale et de l’Amérique du Sud, où il hiverne. Les sites importants pour le rufa se trouvent aux endroits suivants : estuaire du fleuve Nelson, côte de la baie James, archipel de Mingan, baie du Delaware, côte de la Floride; côte du Texas; lac Last Mountain, lac Chaplan/lac Old Wives et lacs Quill; Guyane française; Reentrâncias Maranhenses; Lagoa do Peixe et côte du Rio Grande do Sul; baie San Antonio, péninsule Valdes et estuaire de la baie Blanca; baie Lomas, Rio Grande et baie San Sebastian. L’islandica se reproduit dans les îles Devon, Cornwallis, Bathurst, Byam Martin et Melville et dans toutes les îles situées au nord de ces dernières, ainsi que sur les côtes est et nord du Groenland. L’islandica migre en passant par l’Islande ou le nord de la Norvège pour aller hiverner aux Pays Bas, en Angleterre et dans des parties de la France. Les sites importants pour l’islandica se trouvent aux endroits suivants : Porsangerfjord, en Norvège; Latrabjarg, dans le nord ouest de l’Islande; The Wash, dans le Norfolk, en Angleterre; mer des Wadden, aux Pays Bas; Pertuis Charentais et Bretagne, en France. Il y a chevauchement connu des aires de reproduction du rufa et de l’islandica dans les îles Prince de Galles et Somerset et le nord de l’île de Baffin. Le roselaari se reproduit sur les côtes nord ouest et centre ouest de l’Alaska, et il migre le long de la côte sud de cet État américain, de la côte ouest du Canada et de la côte nord ouest des États Unis. Le roselaari hiverne dans le sud des États Unis et dans l’ensemble de l’Amérique centrale. Les sites importants pour le roselaari se trouvent aux endroits suivants : delta du Yukon Kuskokwim; delta du fleuve Copper; Grays Harbor et baie de Willapa; baie de San Francisco; golfe de Santa Clara et delta du fleuve Colorado; complexe lagunaire Ojo de Liebre.
rufa
En 2012, la population de rufa était estimée à 42 000 individus d'après des relevés printaniers exhaustifs menés sur la côte de l'Atlantique et des travaux réalisés dans le nord-ouest du golfe du Mexique (Andres et al., 2012). Les analyses de la meilleure information accessible tirée des sites d’hivernage et des haltes migratoires laissent entrevoir un déclin constant de la population de rufa pendant les années 2000 (U.S. Fish and Wildlife Service, 2014), suivi d'une période de stabilité potentielle (à un niveau bien inférieur à ceux des années 1980 et 1990) de 2009 à 2014 (Dey et al., 2011; Andres et al., 2012; U.S. Fish and Wildlife Service, 2014).
Au sein de l'aire de reproduction, l'habitat convenable n'est pas continu dans toute l'aire de répartition du rufa, et il semble que ce ne soit pas tout l'habitat convenable potentiel qui est occupé actuellement. Au Nunavut, le rufa niche aux îles Coats et Mansel, dans le nord de la baie d'Hudson, sur l'île Southampton, sur la côte est (Godfrey, 1986) et aux îles du bassin Foxe (p. ex. île Prince Charles, île Rowley et côte ouest de l'île de Baffin [Niles et al., 2005; R.I.G. Morrison, obs. pers.]), de même que probablement jusqu'à l'extrémité ouest de la région de la presqu'île de Boothia, jusqu'à l'île King William (Niles et al., 2005) et jusqu'à l'île Victoria (Parmelee et al., 1967; P. Marra, comm. pers.). Il ne semble pas y avoir d'habitat convenable sur le territoire entre le nord de la baie d'Hudson et le bassin Rasmussen (Niles et al., 2005), et la sous-espèce n'a jamais été observée dans cette région (Godfrey, 1986, 1992) ni dans les basses terres du Rasmussen (Johnston et al., 2000). Bien qu'il semble y avoir de l'habitat convenable à l'île Banks, dans les Territoires du Nord-Ouest, à la limite ouest des îles de l'Arctique, aucune mention de bécasseaux nichant dans cette région n'a été consignée (Manning et al., 1956; V. Johnston, comm. pers., 2005).
Durant la migration vers le nord, de grandes volées de bécasseaux sont observées dans le sud de la baie James à la fin mai ou au début juin (R.I.G. Morrison, données inédites); elles proviennent probablement directement de la baie du Delaware (Delaware et New Jersey, aux États-Unis) (Morrison et Harrington, 1992). Les données obtenues à partir de rufa munis d'un géolocalisateur au Texas laissent deviner une halte migratoire près du fleuve Nelson, sur la côte ouest de la baie d'Hudson, dans le nord du Manitoba, au Canada. Des relevés de suivi ont confirmé la présence de larges concentrations de Bécasseaux maubèches (dénombrement maximal au sol sur une journée de 1 900 individus) à quelque 25 km à l'est de la rivière Hayes, au Manitoba, et des oiseaux ont également été observés dans les secteurs nord et est de l'embouchure du fleuve Nelson (A. McKeller, données inédites). En outre, des oiseaux munis d'un nano-émetteur (transmetteur VHFNote2de bas de page) dans la baie du Delaware ont aussi été détectés lors de ces relevés (A. McKeller, données inédites). De grandes concentrations sont parfois observées autour du lac Ontario, mais il s'agit probablement d'oiseaux détachés du groupe principal de migrateurs à cause de conditions météorologiques défavorables (McRae, 1982; Weir, 1989; Morrison et Harrington, 1992). Toutefois, il est possible que le lac Ontario offre un refuge aux oiseaux migrateurs lorsque les conditions sont difficiles, comme en témoigne l'observation, le 4 juin 2003, au parc provincial Presqu'île, en Ontario, d'un oiseau en piètre condition qui avait été bagué initialement durant la mauvaise saison de ponte de limules en mai 2003 (Niles et al., 2008) (P.M. González, comm. pers., 2015). Plusieurs observations d'oiseaux bagués en Argentine et l'observation au parc provincial Presqu'île d'un oiseau portant une bague colorée posée à la Lagoa do Peixe, dans le sud du Brésil (Clive Goodwin, base de données sur les oiseaux du comté de Northumberland) indiquent que ces oiseaux comprennent des migrateurs de la population de rufa du sud. Plusieurs Bécasseaux maubèches marqués (du Chili, de l'Argentine, du Brésil et de la baie du Delaware) ont été observés ensemble le même jour au lac Saint-Pierre (près de Yamachiche, au Québec) en 2007 (Y. Aubry, comm. pers., 2015).
Durant la migration vers le sud, un grand nombre de bécasseaux passent par la côte sud-ouest de la baie d'Hudson (Manitoba et Ontario) ainsi que par les côtes ouest et sud de la baie James (Ontario) en juillet et en août (Hope et Short, 1944; Manning, 1952; Ross et al., 2003). Le coin sud-est de l'île Akimiski, au Nunavut, semble aussi important pour les bécasseaux. De plus, de grands nombres d'individus ont été signalés le long de la baie de Rupert (sud de la baie James) et de la baie Boatswain (extrémité nord-est de la baie de Rupert, au Québec) (Benoit, 2004). De 2006 à 2015, l'observation dans la réserve de parc national du Canada de l'Archipel-de-Mingan d'un grand nombre d'oiseaux capturés et marqués avec des bagues de couleur au Chili, en Argentine et au Brésil confirme l'identité de ces derniers, c'est-à-dire qu'ils appartiennent à la population de rufa hivernant dans le sud de l'Amérique du Sud ainsi que dans les États de Maranhão et de Ceará dans le nord de l'Amérique du Sud (Y. Aubry, comm. pers., 2015). Ouellet (1969) a identifié 4 bécasseaux prélevés à l’île d’Anticosti dans un groupe de 200 oiseaux comme appartenant à la sous-espèce rufa.
Parmi les zones importantes du rufa se trouvant dans ses voies migratoires, à l'extérieur du Canada, figurent Río Gallegos, la péninsule Valdés, San Antonio Oeste (Patagonie, Argentine) et l'estuaire de la baie Blanca (Buenos Aires, Argentine); la Lagoa do Peixe et l'État côtier de Río Grande do Sul (sud-est du Brésil); Maranhão (nord du Brésil); le Suriname et la Guyane française; le sud-est des États-Unis (p. ex. depuis la Floride jusqu'à la Caroline du Nord); les îles-barrières de la Virgine jusqu'au Massachusetts et la baie du Delaware (González, 2005; Niles et al., 2008; Cohen et al., 2009; Baker et al., 2013; U.S. Fish and Wildlife Service, 2014).
Les principales aires d’hivernage utilisées par le rufa sont réputées englober le centre de la côte floridienne du golfe du Mexique, le sud-est des États-Unis (soit la Géorgie et la Caroline du Sud), le nord-ouest du golfe du Mexique (depuis l'État de Tamaulipas, au Mexique, jusqu'à la Louisiane, en passant par Laguna Madre, au Texas), la côte nord-est du Brésil (soit dans les États de Maranhão et de Ceará), et le littoral atlantique de l'Argentine et du Chili (principalement de la Terre de Feu, qui couvre les deux pays) (Niles et al., 2008; Andres et al., 2012; U.S. Fish and Wildlife, 2014). Le nombre de Bécasseaux maubèches hivernant dans les Caraïbes est inconnu mais, selon des données tirées d'oiseaux munis d'un géolocalisateur, cette région serait un important lieu d'hivernage (U.S. Fish and Wildlife Service, 2014).
roselaari
La tendance démographique du roselaari n'est pas déterminée avec certitude et elle est complexe, car les connaissances relatives à la distribution hivernale de la sous-espèce sont incomplètes (Morrison et al., 2006; Andres et al., 2012). La population mondiale de roselaari était estimée en 2012 à environ 17 000 individus (intervalle de confiance à 95 % fondé sur des mesures statistiques de la précision des données = 14 000-20 000) d'après les résultats des activités de baguage et de marquage-recapture (Andres et al., 2012; Carmona et al., 2013).
Chez le roselaari, on a établi des corrélations évidentes entre les aires d’hivernage dans le nord-ouest du Mexique, les haltes migratoires dans l'État de Washington, aux États-Unis, et les lieux de reproduction dans le nord-ouest et le nord de l'Alaska ainsi qu'à l'île Wrangel, en Russie (Buchanan et al., 2010, 2011; U.S. Fish and Wildlife Service, 2011; Andres et al., 2012; Carmona et al., 2013). De petits nombres de roselaari sont aussi répertoriés en Californie et dans le nord-ouest du golfe du Mexique (Andres et al., 2012). D'après les données de géolocalisation et de réobservation d'oiseaux bagués à ce jour, la quasi-totalité, voire la totalité, des Bécasseaux maubèches hivernant dans le nord-ouest du golfe du Mexique appartiennent à la sous-espèce rufa (U.S. Fish and Wildlife Service, 2014). À la lumière de cette nouvelle information, on croit que la population de roselaari qui fréquente le Canada (le long de la côte du Pacifique en Colombie-Britannique) représente moins de 1 % de la population mondiale actuelle.
islandica
On ne dispose d'aucune nouvelle information sur l'islandica au Canada, mais des données provenant d'aires d'hivernage européennes laissent croire à un faible déclin à court terme (2003-2012) (Andres et al., 2012; Nagy et al., 2014) et à une fluctuation de la tendance à long terme (Nagy et al., 2014). La population d'islandica au Canada était estimée à 80 000 individus (Morrison et al., 2006, 2007; Andres et al., 2012).
Cette sous-espèce hiverne, en Europe, sur le bord de la mer au Royaume-Uni et aux Pays-Bas, et niche dans le nord-est de l'Extrême-Arctique canadien, probablement jusqu'à l'île Prince Patrick (Territoires du Nord-Ouest), vers l'ouest; jusqu'à l'île Prince-de-Galles (Nunavut), vers le sud; le long de la côte nord du Groenland (Manning et Macpherson, 1961; Godfrey, 1992; COSEWIC, 2007). Il faut mener des recherches pour déterminer s'il y a chevauchement entre les aires de reproduction du rufa et de l'islandica (Morrison et Harrington, 1992). La sous-espèce islandica migre vers le nord en passant par l'Islande et le nord de la Norvège.
3.3 Besoins du Bécasseau maubèche
Habitat de reproduction
Le Bécasseau maubèche a besoin d'une toundra sèche, légèrement élevée et exempte de couverture de neige pour nicher. Les nids sont de simples dépressions aménagées dans le sol, souvent dans de petites touffes de végétation (COSEWIC, 2007). Les mâles enlèvent la végétation du site du nid et créent une dépression dans le sol, qu'ils garnissent de lichens et de feuilles mortes. Les nids sont généralement situés à une altitude de moins de 150 m au-dessus du niveau de la mer, à une distance maximale de 50 km de la côte (d'après l'exercice de modélisation du paysage du New Jersey ENSP et de la Rutgers University, dans Niles et al., 2007). Les nids sont isolés dans le paysage, souvent espacés entre 0,75-1 km et 15 km (Niles et al., 2007). Après l'éclosion, les Bécasseaux maubèches ont besoin d'accéder à des milieux d'eau douce abritant des invertébrés dont ils peuvent se nourrir, notamment des insectes (p. ex. larves de moustiques) et d'autres arthropodes (p. ex. araignées) (Harrington, 2001; Niles et al., 2008; U.S. Fish and Wildlife Service, 2014). Les oisillons peuvent se déplacer sur de grandes superficies (plusieurs kilomètres).
Haltes migratoires
Pour se nourrir et se reposer, le Bécasseau maubèche a besoin d'un habitat de qualitéNote3de bas de page (généralement des milieux côtiers marins ou estuariens, mais aussi des lacs salés intérieurs) dans un petit nombre de haltes migratoires importantes. Ces milieux ne servant pas à la reproduction doivent être relativement exempts de perturbations anthropiques; dans les haltes migratoires, l'espèce utilise les plages sablonneuses, flèches littorales, bancs de sable, battures sablonneuses/vaseuses, restingas (c'est-à-dire plateformes rocheuses intertidales érodées par les vagues), plaines rocheuses intertidales, rizières et marais salés (Niles et al., 2007). Pendant la migration printanière dans la baie du Delaware, le rufa a besoin de limules (Limulus polyphemus), en fraye, qui préfèrent les plages dominées par des sédiments sablonneux grossiers (Niles et al., 2007). Lorsque les Bécasseaux maubèches viennent tout juste d'arriver ou sont sur le point de partir (c.-à-d. au début ou à la fin de la migration), ils doivent combler leurs demandes énergétiques sur une courte période; c'est pourquoi une nourriture facile à digérer (soit des proies à coquille mince ou dépourvues de coquille telles que les bivalves juvéniles, les œufs de limules et les vers marins) doit être abondante dans les haltes migratoires (Piersma et al., 1999; van Gils et al., 2005a, 2005b; Niles et al., 2008; Cohen et al., 2011).
Habitat d'hivernage
Les milieux côtiers marins et estuariens utilisés, en période d'hivernage, par le Bécasseau maubèche sont semblables à ceux utilisés pendant la migration (c'est-à-dire aux haltes migratoires). Le Bécasseau maubèche hiverne le long de plages sablonneuses, mais fréquentent aussi les bancs de tourbe, marais salés, lagunes saumâtres, battures de vase, et restingas et plaines rocheuses intertidales. Il doit avoir accès à de la nourriture (naissain de moules et myes, petits crabes, vers polychètes [Baker et al., 2013]). Les habitats d'alimentation et de repos doivent être relativement exempts de perturbations anthropiques.
Habitat des individus immatures avant leur reproduction
On croit que tous les Bécasseaux maubèches immatures demeurent dans les milieux ne servant pas à la reproduction dans leur deuxième été de vie, à des latitudes méridionales, dans des milieux aux caractéristiques possiblement semblables à celles des haltes migratoires et de l'habitat d'hivernage (U.S. Fish and Wildlife Service, 2014). Certains individus de la sous-espèce rufa dans leur deuxième année civile ont été capturés en Argentine, ce qui donne à penser que certains individus immatures peuvent suivre les adultes vers des haltes migratoires post-reproduction situées plus au sud avant de réaliser leur premier vol pré-reproduction avec ces mêmes individus adultes (P.M. González, comm. pers., 2015). De petites bandes de Bécasseaux maubèches immatures ont été observées à la Lagoa do Peixe (Belton, 1984; Serrano, 2001) et sur d'autres plages le long de la côte du Rio Grande do Sul, au Brésil (Scherer et Petry, 2012), à Punta Rasa, en Argentine (Blanco et Carbonell, 2001), sur l'île Margarita, au large du Venezuela (Azpiroz et Rodriguez-Ferraro, 2006) et en d'autres lieux partout dans les Amériques (Baker et al., 2013). Un grand nombre d’oiseaux non nicheurs (présumés être des roselaari) ont été enregistrés de juin à août dans le nord-est du golfe de la Californie, au Mexique (Soto-Montoya et al., 2009).
Facteurs limitatifs
À l'instar de nombreux oiseaux arctiques nichant au sol, le Bécasseau maubèche est limité par une productivité généralement faible, , laquelle peut être quasi nulle certaines années (COSEWIC, 2007; Meltofte et al., 2007; Niles et al., 2008). La productivité est limitée par les conditions météorologiques (p. ex. la fonte des neiges tardive peut entraîner une réduction du nombre d'invertébrés proies, de mauvaises conditions peuvent influer sur la capacité thermorégulatrice des poussins et entraîner une forte mortalité) et l'abondance des prédateurs (généralement associée aux cycles asymétriques des lemmings [Lemmus spp. et Dicrostonyx spp.] survenant à des intervalles de 3-4 ans) (Fraser et al., 2013). L'accès à des haltes migratoires clés pendant la migration printanière peut être un facteur limitatif pour l'espèce. Le Bécasseau maubèche a besoin de ressources alimentaires adéquates pour survivre pendant les longs vols, surmonter les changements physiologiques adaptatifs et prévenir les périodes de pénuries de nourriture dans les lieux de reproduction arctiques (Baker et al. 2004; Morrison 2006; Morrison et al. 2007; Niles et al. 2008; McGowan et al. 2011). Il a aussi besoin de lieux ne servant pas à la reproduction qui sont pourvus de ressources au bon moment de leur cycle vital annuel aux fins de la mue (plumes du corps et plumes de vol). Cela est particulièrement important pour les migrateurs de longue distance, comme la sous espèce rufa (plus précisément les individus de la population de la Terre de Feu), dont la période de mue chevauche celle de la migration (Buehler et Piersma, 2008). Les variations notées quant à l'utilisation de l'habitat, au taux d'alimentation et aux stratégies de migration peuvent aussi être influencées par la présence d'oiseaux de proie (Pomeroy et al., 2006; Niles et al., 2008; Watts, 2009).
4. Menaces
4.1 Évaluation des menaces
La classification des menaces (tableau 2) présentée ci-dessous est fondée sur le système unifié de classification des menaces proposé par l'Union internationale pour la conservation de la nature (IUCN) et le Partenariat pour les mesures de conservation (Conservation Measures Partnership, ou CMP) (IUCN-CMP) et a été modifiée en 2011 d'après l'expérience de son utilisation par le COSEPAC et les équipes de rétablissement. Le présent calculateur des menaces tient compte des normes internationales visant l'identification et l'évaluation des menaces élaborées par la Commission de la sauvegarde des espèces de l'IUCN, le CMP (Salafsky et al., 2008) et The Nature Conservancy. Les normes sont utilisées par le COSEPAC, le programme de conservation et de gestion des oiseaux migrateurs du Service canadien de la faune d'Environnement et Changement climatique Canada, la province de la Colombie-Britannique et NatureServe. Leur utilisation dans la planification du rétablissement aux termes de la LEP est sur le point d'être adoptée en prévision de l'échange et de la coordination améliorés des données sur les espèces en péril tant au sein du gouvernement fédéral qu'entre les gouvernements fédéral, provinciaux et territoriaux, lesquels ont également adopté le système.
| Catégorie de la menace | # | Menace | Sous-espèce | Impactf | Portéeg | Gravitéh | Immédiatetéi |
|---|---|---|---|---|---|---|---|
| 1 Développement résidentiel et commercial | 1.1 | Zones résidentielles et urbaines | rufa | Faible | Restreinte | Légère | Élevée |
| 1 Développement résidentiel et commercial | 1.1 | Zones résidentielles et urbaines | roselaari | Faible | Restreinte | Légère | Élevée |
| 1 Développement résidentiel et commercial | 1.1 | Zones résidentielles et urbaines | islandica | Faible | Restreinte | Légère | Élevée |
| 1 Développement résidentiel et commercial | 1.2 | Zones commerciales et industrielles | rufa | Faible | Restreinte-petite | Légère | Élevée |
| 1 Développement résidentiel et commercial | 1.2 | Zones commerciales et industrielles | roselaari | Faible | Restreinte | Légère | Élevée |
| 1 Développement résidentiel et commercial | 1.2 | Zones commerciales et industrielles | islandica | Faible | Restreinte | Légère | Modérée |
| 1 Développement résidentiel et commercial | 1.3 | Zones touristiques et récréatives | rufa | Inconnu | Inconnue | Inconnue | Élevée |
| 1 Développement résidentiel et commercial | 1.3 | Zones touristiques et récréatives | roselaari | - | - | - | - |
| 1 Développement résidentiel et commercial | 1.3 | Zones touristiques et récréatives | islandica | - | - | - | - |
| 2 Agriculture et aquaculture | 2.1 | Cultures annuelles et pérennes de produits autres que le bois | rufa | Inconnu | Inconnue | Inconnue | Élevée |
| 2 Agriculture et aquaculture | 2.1 | Cultures annuelles et pérennes de produits autres que le bois | roselaari | - | - | - | - |
| 2 Agriculture et aquaculture | 2.1 | Cultures annuelles et pérennes de produits autres que le bois | islandica | - | - | - | - |
| 2 Agriculture et aquaculture | 2.3 | Élevage de bétail | rufa | Inconnu | Inconnue | Inconnue | Élevée |
| 2 Agriculture et aquaculture | 2.3 | Élevage de bétail | roselaari | - | - | - | - |
| 2 Agriculture et aquaculture | 2.3 | Élevage de bétail | islandica | - | - | - | - |
| 2 Agriculture et aquaculture | 2.4 | Aquaculture en mer et en eau douce | rufa | Inconnu | Restreinte | Inconnue | Élevée |
| 2 Agriculture et aquaculture | 2.4 | Aquaculture en mer et en eau douce | roselaari | - | - | - | - |
| 2 Agriculture et aquaculture | 2.4 | Aquaculture en mer et en eau douce | islandica | - | - | - | - |
| 3 Production d'énergie et exploitation minière | 3.1 | Forage pétrolier et gazier | rufa | - | - | - | - |
| 3 Production d'énergie et exploitation minière | 3.1 | Forage pétrolier et gazier | roselaari | Faible | Petite | Légère | Élevée |
| 3 Production d'énergie et exploitation minière | 3.1 | Forage pétrolier et gazier | islandica | - | - | - | - |
| 3 Production d'énergie et exploitation minière | 3.2 | Exploitation de mines et de carrières | rufa | Faible | Petite | Légère | Élevée |
| 3 Production d'énergie et exploitation minière | 3.2 | Exploitation de mines et de carrières | roselaari | - | - | - | - |
| 3 Production d'énergie et exploitation minière | 3.2 | Exploitation de mines et de carrières | islandica | - | - | - | - |
| 3 Production d'énergie et exploitation minière | 3.3 | Énergie renouvelable | rufa | Faible | Petite | Légère | Élevée |
| 3 Production d'énergie et exploitation minière | 3.3 | Énergie renouvelable | roselaari | - | - | - | - |
| 3 Production d'énergie et exploitation minière | 3.3 | Énergie renouvelable | islandica | - | - | - | - |
| 4 Corridors de transport et de service | 4.3 | Voies de transport par eau | rufa | Négligeable | Négligeable | Légère | Modérée |
| 4 Corridors de transport et de service | 4.3 | Voies de transport par eau | roselaari | Négligeable | Négligeable | Légère | Modérée |
| 4 Corridors de transport et de service | 4.3 | Voies de transport par eau | islandica | Négligeable | Négligeable | Négligeable | Faible |
| 5 Utilisation des ressources biologiques | 5.1 | Chasse et capture d'animaux terrestres | rufa | Inconnu | Restreinte | Inconnue | Inconnue |
| 5 Utilisation des ressources biologiques | 5.1 | Chasse et capture d'animaux terrestres | roselaari | - | - | - | - |
| 5 Utilisation des ressources biologiques | 5.1 | Chasse et capture d'animaux terrestres | islandica | Faible | Petite | Légère | Élevée |
| 5 Utilisation des ressources biologiques | 5.4 | Pêche et récolte de ressources aquatiques | rufa | Moyen | Généralisée | Légère | Élevée |
| 5 Utilisation des ressources biologiques | 5.4 | Pêche et récolte de ressources aquatiques | roselaari | - | - | - | - |
| 5 Utilisation des ressources biologiques | 5.4 | Pêche et récolte de ressources aquatiques | islandica | Faible | Petite | Légère | Modérée |
| 6 Intrusions et perturbations humaines | 6.1 | Activités récréatives | rufa | Faible | Généralisée | Légère | Élevée |
| 6 Intrusions et perturbations humaines | 6.1 | Activités récréatives | roselaari | Faible | Grande | Légère | Élevée |
| 6 Intrusions et perturbations humaines | 6.1 | Activités récréatives | islandica | - | - | - | - |
| 6 Intrusions et perturbations humaines | 6.3 | Travail et autres activités | rufa | Négligeable | Grande | Négligeable | Élevée |
| 6 Intrusions et perturbations humaines | 6.3 | Travail et autres activités | roselaari | - | - | - | - |
| 6 Intrusions et perturbations humaines | 6.3 | Travail et autres activités | islandica | - | - | - | - |
| 7 Modifications des systèmes naturels | 7.2 | Gestion et utilisation de l'eau et exploitation de barrages | rufa | Inconnu | Restreinte | Inconnue | Élevée |
| 7 Modifications des systèmes naturels | 7.2 | Gestion et utilisation de l'eau et exploitation de barrages | roselaari | - | - | - | - |
| 7 Modifications des systèmes naturels | 7.2 | Gestion et utilisation de l'eau et exploitation de barrages | islandica | - | - | - | - |
| 7 Modifications des systèmes naturels | 7.3 | Autres modifications de l'écosystème | rufa | Inconnu | Grande | Inconnue | Élevée |
| 7 Modifications des systèmes naturels | 7.3 | Autres modifications de l'écosystème | roselaari | Inconnu | Grande | Inconnue | Élevée |
| 7 Modifications des systèmes naturels | 7.3 | Autres modifications de l'écosystème | islandica | - | - | - | - |
| 8 Espèces et gènes envahissants ou autrement problématiques | 8.1 | Espèces exotiques (non indigènes) envahissantes | rufa | Faible | Petite | Légère | Élevée |
| 8 Espèces et gènes envahissants ou autrement problématiques | 8.1 | Espèces exotiques (non indigènes) envahissantes | roselaari | Faible | Grande | Légère | Élevée |
| 8 Espèces et gènes envahissants ou autrement problématiques | 8.1 | Espèces exotiques (non indigènes) envahissantes | islandica | - | - | - | - |
| 8 Espèces et gènes envahissants ou autrement problématiques | 8.2 | Espèces indigènes problématiques | rufa | Faible | Généralisée | Légère | Élevée |
| 8 Espèces et gènes envahissants ou autrement problématiques | 8.2 | Espèces indigènes problématiques | roselaari | Inconnu | Généralisée | Inconnue | Élevée |
| 8 Espèces et gènes envahissants ou autrement problématiques | 8.2 | Espèces indigènes problématiques | islandica | - | - | - | - |
| 9 Pollution | 9.1 | Eaux usées domestiques et urbaines | rufa | Inconnu | Inconnue | Inconnue | Élevée |
| 9 Pollution | 9.1 | Eaux usées domestiques et urbaines | roselaari | Faible | Grande | Légère | Modérée |
| 9 Pollution | 9.1 | Eaux usées domestiques et urbaines | islandica | - | - | - | - |
| 9 Pollution | 9.2 | Effluents industriels et militaires | rufa | Élevé-Moyen | Grande | Élevée-modérée | Modérée |
| 9 Pollution | 9.2 | Effluents industriels et militaires | roselaari | - | - | - | - |
| 9 Pollution | 9.2 | Effluents industriels et militaires | islandica | Négligeable | Restreinte | Négligeable | Modérée |
| 9 Pollution | 9.3 | Effluents agricoles et sylvicoles | rufa | Négligeable | Petite | Négligeable | Élevée |
| 9 Pollution | 9.3 | Effluents agricoles et sylvicoles | roselaari | Faible | Grande | Légère | Modérée |
| 9 Pollution | 9.3 | Effluents agricoles et sylvicoles | islandica | - | - | - | - |
| 9 Pollution | 9.4 | Déchets solides et ordures | rufa | Inconnu | Inconnue | Légère | Élevée |
| 9 Pollution | 9.4 | Déchets solides et ordures | roselaari | - | - | - | - |
| 9 Pollution | 9.4 | Déchets solides et ordures | islandica | - | - | - | - |
| 11 Changements climatiques et phénomènes météorologiques violents | 11.1 | Déplacement et altération de l'habitat | rufa | Non calculéj | Généralisé | Inconnue | Faible |
| 11 Changements climatiques et phénomènes météorologiques violents | 11.1 | Déplacement et altération de l'habitat | roselaari | Non calculéj | Grande | Inconnue | Faible |
| 11 Changements climatiques et phénomènes météorologiques violents | 11.1 | Déplacement et altération de l'habitat | islandica | Non calculéj | Généralisée | Inconnue | Faible |
| 11 Changements climatiques et phénomènes météorologiques violents | 11.4 | Tempêtes et inondations | rufa | Inconnu | Généralisée | Inconnue | Modérée |
| 11 Changements climatiques et phénomènes météorologiques violents | 11.4 | Tempêtes et inondations | roselaari | Inconnu | Grande | Inconnue | Modérée |
| 11 Changements climatiques et phénomènes météorologiques violents | 11.4 | Tempêtes et inondations | islandica | - | - | - | - |
f Impact – Mesure dans laquelle on observe, infère ou soupçonne que l'espèce est directement ou indirectement menacée dans la zone d'intérêt. Le calcul de l'impact de chaque menace est fondé sur sa gravité et sa portée et prend uniquement en compte les menaces présentes et futures. L'impact d'une menace est établi en fonction de la réduction de la population de l'espèce, ou de la diminution dégradation de la superficie d'un écosystème. Le taux médian de réduction de la population ou de la superficie pour chaque combinaison de portée et de gravité correspond aux catégories d'impact suivantes : très élevé (déclin de 75 %), élevé (40 %), moyen (15 %) et faible (3 %). Inconnu : catégorie utilisée quand l'impact ne peut être déterminé (p. ex. lorsque les valeurs de la portée ou de la gravité sont inconnues); non calculé : l'impact n'est pas calculé lorsque la menace se situe en dehors de la période d'évaluation (p. ex. l'immédiateté est non significative/négligeable ou faible puisque la menace n'existait que dans le passé); négligeable : lorsque la valeur de la portée ou de la gravité est négligeable; n'est pas une menace : lorsque la valeur de la gravité est neutre ou qu'il y a un avantage possible.
g Portée – Proportion de l'espèce qui, selon toute vraisemblance, devrait être touchée par la menace d'ici 10 ans. Correspond habituellement à la proportion de la population de l'espèce dans la zone d'intérêt (généralisée = 71-100 %; grande = 31-70 %; restreinte = 11-30 %; petite = 1-10 %; négligeable < 1 %).
h Gravité – Au sein de la portée, niveau de dommage (habituellement mesuré comme l'ampleur de la réduction de la population) que causera vraisemblablement la menace sur l'espèce d'ici une période de 10 ans ou de 3 générations (extrême = 71-100 %; élevée = 31-70 %; modérée = 11-30 %; légère = 1-10 %; négligeable < 1 %; neutre ou avantage possible ≥ 0 %).
i Immédiateté – Élevée = menace toujours présente; modérée = menace pouvant se manifester uniquement dans le futur (à court terme [< 10 ans ou 3 générations]) ou pour l'instant absente (mais susceptible de se manifester de nouveau à court terme); faible = menace pouvant se manifester uniquement dans le futur (à long terme) ou pour l'instant absente (mais susceptible de se manifester de nouveau à long terme); non significative/négligeable = menace qui s'est manifestée dans le passé et qui est peu susceptible de se manifester de nouveau, ou menace qui n'aurait aucun effet direct, mais qui pourrait être limitative.
j = en dehors de la période d'évaluation
4.2 Description des menaces
Les rufa et islandica nicheurs font actuellement face à peu de menaces durant leur court séjour, alors qu'ils sont largement dispersés dans leur vaste aire de reproduction de l'Arctique canadien. Les menaces (telles qu'indiquées dans le tableau 2), effluents industriels et militaires ainsi que changements climatiques et phénomènes météorologiques violents, constituent des exceptions notables. Les rufa et roselaari concentrés dans les haltes migratoires canadiennes sont exposés à un certain nombre de menaces (telles qu'indiquées dans le tableau 2) qui sont considérées comme ayant le plus grand impact sur les populations lors de leur passage au Canada, soient pêche et récolte de ressources aquatiques, activités récréatives, effluents industriels et militaires et changements climatiques et phénomènes météorologiques violents.
Les menaces d'impact faible à élevé sont énumérées conformément au tableau d'évaluation du calculateur de menaces (tableau 2) ci-dessus et sont décrites en détail ci-dessous.
1. Développement résidentiel et commercial
1.1 Zones résidentielles et urbaines et 1.2 Zones commerciales et industrielles (rufa, roselaari et islandica) :
La population humaine continue de croître, et cette croissance, combinée au désir de vivre en milieu côtier, crée des conflits à mesure que les humains construisent à l'intérieur ou à proximité des habitats privilégiés par les Bécasseaux maubèches. Le long de la côte de l'Atlantique aux États-Unis, près de un tiers du littoral peut encore être développé. Le régime foncier de certains emplacements procure une certaine protection de l'habitat (p. ex. organisation fédérale, d'un État ou privée vouée à la conservation des terres, ou servitude de conservation permanente) (U.S. Fish and Wildlife Service, 2014). En Amérique du Sud, le développement urbain, commercial et industriel peut poser un risque pour le rufa le long de la côte nord-est du Brésil et en Argentine (p. ex. Río Gallegos et portions argentines de la Terre de Feu) (U.S. Fish and Wildlife Service, 2014; WHSRN, 2015). L'empiètement et la transformation des battures et des marais salés à des fins de développement urbain, commercial et industriel sont préoccupants pour les oiseaux de rivage, à mesure que la ville de Río Gallegos, en Argentine, s'étend vers la côte (Ferrari et al., 2002). Près de 10 % de la population d'islandica hiverne le long de la côte atlantique de la France (Bocher et al., 2012), où l'habitat de repos convenable est peut-être limité à cause des pressions exercées par les perturbations et le développement urbain, commercial et industriel (Leyrer et al., 2014).
1.3 Zones touristiques et récréatives (rufa) :
Des installations touristiques et des points d'accès continuent d'être aménagés le long de la plage de la halte migratoire de l'aire naturelle protégée de la baie San Antonio, en Argentine, dans le cadre d'un nouveau plan d'urbanisme. Cette expansion pourrait causer la dégradation de l'habitat des oiseaux de rivage (WHSRN, 2015). Les aires récréatives constituent probablement une menace localisée pour le Bécasseau maubèche au sein de ses aires de migration et d'hivernage, mais l'étendue et l’impact de cette menace ne sont pas connus.
2. Agriculture et aquaculture
2.1 Cultures annuelles et pérennes de produits autres que le bois (rufa) :
Les haltes migratoires au Brésil peuvent être touchées défavorablement par les pratiques agricoles adjacentes qui altèrent les propriétés hydrologiques et augmentent l’envasement d’importants habitats lagunaires (Niles et al., 2008; U.S. Fish and Wildlife Service, 2014). Les milieux côtiers secs adjacents, près de la Lagoa do Peixe, au Brésil, et de Río Gallegos, en Argentine, montrent des signes de dégradation découlant des cultures destinées à l'alimentation (p. ex. oignon, riz, maïs) (U.S. Fish and Wildlife Service, 2014; WHSRN, 2015).
2.3 Élevage de bétail (rufa) :
En Amérique du Sud, l'élevage de bétail est pratiqué sur des terres adjacentes à des réserves, à Río Gallegos, en Argentine (Niles et al., 2008), et le pâturage extensif par le bétail perturbe les milieux côtiers près de la Lagoa do Peixe, sur la côte est du Brésil (WHSRN, 2015).
2.4 Aquaculture en mer et en eau douce (rufa) :
Au Canada, la mytiliculture (dans le cadre de laquelle de jeunes myes sont récoltées par filtration de sable, puis transplantées dans des étendues de sable servant d'aires de croissance voisines) influe sur la qualité de l'habitat d'alimentation du rufa au Québec (Y. Aubry, comm. pers., 2015). L’élevage de crevettes, de même que la perte et la dégradation de l'habitat qui s'ensuivent, a probablement touché le Bécasseau maubèche dans le nord-est du Brésil ces 20 à 25 dernières années (Carlos et al., 2010). La récolte des algues et l'aquaculture dégradent potentiellement la qualité de l'habitat du Bécasseau maubèche en Argentine et à l'île Chiloé, au Chili (U.S. Fish and Wildlife Service, 2014).
3. Production d'énergie et exploitation minière
3.1 Forage pétrolier et gazier (roselaari) :
Le développement de l'industrie pétrolière et gazière, y compris les infrastructures associées, pourrait avoir des impacts considérables sur l'habitat dans le nord de l'Alaska (Alaska Shorebird Group, 2008). Une hausse de la production pétrolière est anticipée en Alaska pour la période 2015-2017, et de nouvelles découvertes en milieu terrestre sont attendues en Arctique (Resource Development Council, 2015).
3.2 Exploitation de mines et de carrières (rufa) :
Les activités minières accrues (p. ex. aux fins de l'extraction de diamants, de minerai de fer, de charbon et d'agrégats), y compris les infrastructures associées en Arctique peuvent menacer les Bécasseaux maubèches. Des mines et des carrières sont aussi exploitées le long des cours d'eau qui traversent les haltes migratoires sur les côtes est (Québec) et ouest (Ontario) de la baie James, et des activités d'exploration sont en cours dans cette région (V. Brownell et autres, comm. pers., 2015). Une montée du prix de l'or a entraîné une hausse des activités d'extraction de ce métal à petite échelle en Amérique du Sud. L'exploitation minière peut directement endommager le lit et les berges des cours d'eau, causer un envasement en aval et libérer dans l'environnement du mercure qui pourrait atteindre la côte par l'intermédiaire des cours d'eau (Alvarez-Berríos et Aide, 2015).
3.3 Énergie renouvelable (rufa) :
Les éoliennes ont des répercussions à la fois directes (c'est-à-dire mortalité due aux collisions) et indirectes (p. ex. perte d'habitat, comportement d'évitement) sur les oiseaux. L'exploitation de l'énergie éolienne est proposée à l'intérieur de l'aire de migration du Bécasseau maubèche au Canada (p. ex. sud-ouest de l'Ontario et berge du lac Ontario) et aux États-Unis, et des éoliennes terrestres sont déjà établies. La croissance de l'industrie de l'énergie éolienne devrait résulter (Zimmerling et al., 2013) d'un effort visant à réduire la pollution au carbone (Executive Office of the President, 2013). On s'attend à une augmentation de l'utilisation de l'énergie éolienne par l'industrie et les communautés dans l'Arctique canadien (M. Lamont, comm. pers., 2015). Depuis 2009, l'exploitation de l'énergie éolienne en tant que source d'électricité a rapidement augmenté au Brésil (Brazil Wind Power, 2015), et l'intérêt pour l'aménagement éolien, surtout en milieu extracôtier, est grandissant (RECHARGE, 2015). Des parcs éoliens sont en fonction sur la côte, dans le nord du Brésil (R.I.G. Morrison, comm. pers., 2015), et l'impact sur le Bécasseau maubèche de ces parcs ainsi que des aménagements éoliens futurs sont inconnus. Il est certain que l'impact environnemental de tels aménagements est évident. Par exemple, les effets environnementaux d'un parc éolien côtier se trouvant dans le nord-est de l'État de Ceará situé à proximité (adjacent à la collectivité de Xavier) était grave (p. ex. prélèvement de grandes quantités de sable, lequel a été remplacé par du sable et de l'argile de carrières, effets sur le transport des sédiments, remblayage de lacs interdunaux, compactage du sol et du sable) (Meireles et al., 2013). L'impact de ce parc éolien côtier sur les espèces sauvages n’est pas clair.
5. Utilisation des ressources biologiques
5.1 Chasse et capture d'animaux terrestres (rufa et islandica) :
La chasse aux oiseaux de rivage, dont les bécasseaux, est pratiquée dans certaines régions, notamment dans les Caraïbes et le centre-nord du Brésil (Harrington, 2001) bien que, dans cette dernière région, cette activité semble avoir diminué grandement au cours de la dernière décennie (Niles et al., 2005). Le Bécasseau maubèche a récemment été ajouté à la liste des espèces qu'il est interdit de chasser en Guyane française (2014) (A. Duncan, comm. pers., 2015), en Guadeloupe (2012) et en Martinique (2013) (Sorenson et Douglas, 2013). La chasse (p. ex. de subsistance et récréative, à la fois légale et illégale) est encore courante dans les Guyanes et les Caraïbes, le long de la côte nord de l'Amérique du Sud et peut-être dans d'autres territoires. Les oiseaux hivernant dans le sud qui pourraient fréquenter ces emplacements pendant la migration et/ou pendant des phénomènes météorologiques sont potentiellement à risque, et une évaluation de cette menace est donc nécessaire. La sous-espèce islandica est encore une espèce chassable en France (Bocher et al., 2012; A. Duncan, comm. pers., 2015), mais le gouvernement envisage de retirer le Bécasseau maubèche de la liste des espèces chassées (Sorenson et Douglas, 2013; A. Duncan, comm. pers., 2015).
5.4 Pêche et récolte de ressources aquatiques (rufa et islandica) :
Le principal facteur de causalité connu du déclin de la population de rufa à son halte migratoire dans la baie du Delaware était la récolte commerciale de limules pratiquée à leur halte migratoire finale. Plusieurs études confirment que les œufs de limules composent principalement le régime alimentaire des bécasseaux et d'autres oiseaux de rivage dans la baie du Delaware pendant la migration vers le nord (Morrison et Harrington, 1992; Castro et Myers, 1993; Botton et al., 1994; Harrington, 1996, 2001; Tsipoura et Burger, 1999; Haramis et al., 2002, 2007; Clark et al.,2009). Cette source de nourriture autrefois surabondante a été décimée par la surpêche des limules, et on a établi une nette corrélation entre le déclin du rufa et la récolte des limules (U.S. Fish et Wildlife Service, 2014). À mesure que le nombre de limules reproducteurs diminuait, la densité des œufs dans la couche supérieure de sable (5 premiers cm) des plages du New Jersey baissait également. De plus, dans leurs études, Hernandez (2005) et Stillman et al. (2005) ont montré que la densité des œufs était trop faible pour permettre aux bécasseaux de s'alimenter de manière efficiente et de répondre à leurs besoins énergétiques pendant leur halte. Les oiseaux étaient incapables d'atteindre la masse corporelle requise en vue du vol vers les lieux de reproduction arctiques, du moins certaines années (Baker et al., 2004). La récolte des limules dans la baie du Delaware fait aujourd'hui l'objet d'une gestion adaptative, et les restrictions ont permis d'assurer une stabilité apparente de la population de ce crabe (Atlantic States Marine Fisheries Commission, 2015). Aux haltes migratoires utilisées pendant la migration automnale à Cacouna, au Québec, la récolte des algues qui est pratiquée a certaines incidences sur l'habitat de halte migratoire du rufa (Y. Aubry, comm. pers., 2015). En France, certains individus de la sous-espèce islandica peuvent être dérangés par les cueilleurs professionnels de myes ou de coques dans les baies estuariennes pendant l'hiver (Bocher et al., 2012).
6. Intrusions et perturbations humaines
6.1 Activités récréatives (rufa et roselaari) :
De nombreuses études montrent que les perturbations humaines répétées (p. ex. les activités des marcheurs, pêcheurs/collectionneurs, chiens, véhicules hors route, bateaux) peuvent avoir des incidences négatives sur les oiseaux de rivage en altérant leur profil comportemental et leur bilan énergétique (voir par exemple Davidson et Rothwell, 1993; West et al., 2002).
La perturbation est une préoccupation pour le rufa durant sa migration automnale aux Îles-de-la-Madeleine, au Québec. La majorité des activités perturbatrices sont liées à la pêche récréative des coques, aux chars à voile, à l'observation d'espèces sauvages et à l'utilisation de véhicules hors route dans les zones intertidales (ECCC-SCF, Région du Québec, données inédites). La perturbation des oiseaux de rivage a déjà été un grave problème dans la baie du Delaware pendant la migration printanière (Burger et al., 1995; Sitters, 2001), mais la fermeture d'importantes sections du littoral du New Jersey depuis 2003, qui interdit l'accès pendant le pic de la migration, a réussi à réduire efficacement le problème (Burger et al., 2004; Niles et al., 2005). Dans d'autres portions de l'aire de répartition, la perturbation peut constituer un facteur important de dérangement; les oiseaux de rivage étant poussés à abandonner les habitats de choix pour l'alimentation et le repos (U.S. Fish and Wildlife Service, 2014). La perturbation par les humains et les chiens des bandes d'oiseaux qui se reposent ou se nourrissent a été rapportée en Floride, en Géorgie, en Caroline du Nord, en Caroline du Sud, en Virginie et au Massachusetts (Niles et al., 2005). Dans les lieux d’hivernage de la Terre de Feu, les bandes d'oiseaux se reposant à Río Grande sont fréquemment dérangées par les marcheurs, les coureurs, les pêcheurs, les chiens, les véhicules hors route et les motocyclistes (Niles et al., 2005; R.I.G. Morrison, observation personnelle). En Argentine, une perturbation similaire des bécasseaux en migration a été signalée à Río Gallegos, sur la péninsule Valdés, à San Antonio Oeste, dans l'estuaire de la baie Blanca et dans la baie Samborombón (Niles et al., 2005 (P.M. González, comm. pers., 2015). On en connaît peu sur la menace que présentent les perturbations anthropiques pour le roselaari. Toutefois, des haltes migratoires se trouvent à proximité de zones urbaines où les perturbations anthropiques découlant des activités récréatives peuvent exister (G. Donaldson, comm. pers., 2015).
7. Modifications des systèmes naturels
7.2 Gestion et utilisation de l'eau et exploitation de barrages (rufa) :
Nombre de milieux humides importants fréquentés par les oiseaux de rivage migrateurs font l’objet de scénarios de gestion de l’eau dans les Prairies canadiennes (C.L. Gratto-Trevor, comm. pers., 2015), et une telle gestion peut avoir des effets néfastes sur les sources de nourriture et l’habitat de repos convenable pour les oiseaux de rivage migrateurs. La gestion de l’eau (abaissements de l’eau et inondations dans un complexe de milieux humides) dans certains emplacements peut être bénéfique aux oiseaux de rivage si la période et la durée de gestion sont adéquates (Skagen et Thompson, 2013). L’assèchement non réglementé et sans permis des milieux humides est considéré comme une menace pour l’habitat des oiseaux de rivage aux lacs Quill, en Saskatchewan (WHSRN, 2015), tandis que le remblayage est également documenté comme étant une menace pour les milieux humides intérieurs éphémères et temporaires qui sont importants pour les oiseaux de rivage (Skagen et Thompson, 2013). L’altération de l’écoulement de l’eau douce pourrait représenter un des facteurs de stress les plus courants dans les estuaires, les lagunes et les deltas (Sklar et Browder, 1998), perturbant potentiellement les concentrations de nutriments, la salinité, la sédimentation, la topographie, les concentrations d’oxygène dissous et d’autres composantes de l’écosystème. La réponse des écosystèmes à l’altération de l’écoulement de l’eau douce est toutefois complexe et souvent imprévisible (Sklar et Browder, 1998).
7.3 Autres modifications de l'écosystème (rufa et roselaari) :
La majorité du littoral déjà aménagé des États-Unis faisant partie de l'aire de répartition du rufa a subi une certaine forme de stabilisation des berges (p. ex. aménagement de structures rigides telles que les épis, les murs de protection et les brise-lames; aménagement de structures souples telles que les géotubes, les tapis de fibres de coco, les sacs de sable; remblayage des plages (c'est-à-dire l'ajout de sable à une berge qui s'érode pour élargir la plage existante)) (U.S. Fish and Wildlife Service, 2014). Des mesures de stabilisation des berges ont aussi des incidences sur les sites côtiers du Canada (Atlantic Climate Adaptation Solutions, 2011). Les travaux de stabilisation des berges peuvent également être une menace pour le roselaari dans l'ensemble de son aire de répartition (U.S. Fish and Wildlife Service, 2011). La perte des plages et des milieux intertidaux dont a besoin le Bécasseau maubèche s'accélère quand des projets de stabilisation des berges bloquent le déplacement naturel des berges vers les terres, et altèrent la morphologie des plages, la qualité des sédiments et la dynamique de l'eau (voir par exemple Najjar et al., 2000). Les phénomènes météorologiques violents (Lathrop et al., 2013) et la stabilisation des berges avec des structures rigides (Myers, 1986; Jackson et al., 2010) sont également réputés pour dégrader l’habitat requis par les limules en fraye. À mesure que les zones côtières sont aménagées et que le niveau de la mer monte, on s’attend à ce qu’il y ait de plus en plus de tentatives de stabilisation du rivage, ce qui pourrait avoir un impact négatif sur les oiseaux de rivage migrateurs ou hivernants. Il faut continuellement remblayer les plages pour les maintenir, ce qui peut perturber les oiseaux de rivage si les travaux sont effectués alors que les oiseaux s’y trouvent. Le remblayage peut également altérer de manière provisoire et/ou permanente la disponibilité des invertébrés proies pour les oiseaux de rivage (Schlacher et al., 2012; Peterson et al., 2014; U.S. Fish and Wildlife Service, 2014), surtout si les sédiments ajoutés sont trop différents des sédiments naturels. Le rétablissement des invertébrés après les travaux de remblayage dépend de nombreux facteurs, et des incertitudes entourent encore les effets du remblayage sur la communauté des invertébrés et, par le fait même, sur le Bécasseau maubèche (U.S. Fish and Wildlife Service, 2014). Toutefois, le remblayage des plages a aussi le potentiel de mettre en valeur, de restaurer et de créer de l’habitat convenable pour les limules en fraye dans les sites dégradés. Des activités de restauration en cours dans des secteurs clés de la baie du Delaware visent à maintenir à la fois l’habitat du limule et l’habitat des oiseaux de rivage qui dépendent des œufs de limules pour faire le plein d’énergie avant de poursuivre leur migration vers le nord (Siok et Wilson, 2011; Niles et al., 2013a, 2013b; U.S. Fish and Wildlife Service, 2014).
8. Espèces et gènes envahissants ou autrement problématiques
8.1 Espèces exotiques (non indigènes) envahissantes (rufa et roselaari) :
Dans les habitats ne servant pas à la reproduction, le Bécasseau maubèche préfère la végétation clairsemée et nécessite des milieux ouverts et exempts de hauts perchoirs (afin d'éviter les prédateurs). Les plantes envahissantes qui sont ligneuses ou qui forment des touffes ou des tapis denses (p. ex. spartine alterniflore [Spartina alterniflora] peuvent modifier les communautés végétales et nuire à l'habitat des oiseaux de rivage (Niles et al., 2008; U.S. Fish and Wildlife Service, 2014).
8.2 Espèces indigènes problématiques (rufa et roselaari) :
Ces 30 dernières années, les oiseaux de rivage profitent de ce que Butler et al. (2003) ont désigné comme étant un « vide de prédateurs » causé par l'appauvrissement marqué des populations d'oiseaux de proie dû à la persécution et à l'empoisonnement par les pesticides. On ne sait pas très bien si l'augmentation de la prédation par les oiseaux de proie touche les bécasseaux en particulier, mais elle peut être, en général, une source importante de mortalité chez les oiseaux de rivage dans les sites clés (Piersma et al., 1993). Le risque de mortalité directe dans les sites ne servant pas à la reproduction est sans doute faible, mais le risque de prédation peut nuire indirectement aux bécasseaux en les perturbant, en réduisant les épisodes d'alimentation, en restreignant l'accès aux sites d'alimentation de choix et en modifiant le comportement migratoire (voir par exemple Stillman et al., 2005; Pomeroy et al., 2006; Niles et al., 2008). Un épisode de mortalité directe important présumément lié à des proliférations d'algues toxiques (parfois appelées « marées rouges ») a été documenté dans le cas du rufa en Uruguay en 2007 et deux épisodes de mortalité se sont produits dans le sud du Brésil (en 1997 et en 2000) (Buehler et al., 2010). Les bivalves et autres proies privilégiées par les bécasseaux peuvent accumuler des toxines algales s'ils y sont exposés (U.S. Fish and Wildlife Service, 2014). De telles toxines ont été détectées dans des proies se trouvant à l'intérieur des aires ne servant pas à la reproduction du Bécasseau maubèche (Bricelj et al., 2012) et les proliférations d'algues toxiques peuvent donc contribuer à la mortalité du Bécasseau maubèche dans les aires ne servant pas à la reproduction, où le climat est chaud.
9. Pollution
9.1 Eaux usées domestiques et urbaines (rufa et roselaari) :
Jusqu'en 2012, des eaux usées non traitées étaient rejetées dans l'habitat du Bécasseau maubèche à Río Gallegos (U.S. Fish and Wildlife Service, 2014; WHSRN, 2015). Les effets à court et à long terme des eaux usées déjà rejetées sont inconnus. En raison de la proximité du roselaari avec les zones urbaines pendant la période de migration et l'hiver, il est présumé qu'il fréquente des zones potentiellement touchées par les eaux usées et les eaux d'égout (G. Donaldson, comm. pers., 2015).
9.2 Effluents industriels et militaires (rufa) :
Le transport maritime a lieu le long des côtes est (Québec) et ouest (Ontario) de la baie James et dans l'Arctique canadien, et on s'attend à ce que les activités du transport maritime continuent d'augmenter étant donné la prolongation de la période sans glace (Smith et Stephenson, 2013; Pizzolato et al., 2014), ce qui augmente le risque dans cette région. Dans le cas d'un déversement majeur dans une région éloignée, la vitesse de réaction pourrait être inadéquate (MPO, 2012). En Amérique du Nord, d'importantes zones estuariennes, comme la baie du Delaware, les côtes est (Québec) et ouest (Ontario) de la baie James et le golfe du Saint-Laurent, sont exposées à des risques de pollution et à des accidents de transport maritime. Les oiseaux (p. ex. Leighton, 1991; Peterson et al., 2003; Henkel et al., 2012) comme leurs proies invertébrées marines (Blackburn et al., 2014) sont exposés aux produits pétroliers qui se trouvent dans les milieux intertidaux contaminés. Les contaminants environnementaux (hydrocarbures pétroliers, métaux lourds, pesticides et diphényles polychlorés [BPC]) provenant des anciens postes radars du réseau Mid-Canada situés dans le nord de l'Ontario représentent aussi des préoccupations pour les espèces sauvages qui utilisent des sites à proximité. Cependant, des activités d'assainissement sont en cours ou achevées dans la plupart des sites de l'Ontario (Abraham et McKinnon, 2011), et on a constaté que les concentrations de BPC diminuaient à la suite de la suppression de leur principale source terrestre (Abraham et McKinnon, 2011). Les espèces sauvages dans l'archipel de Mingan ou à proximité de celui-ci, dans le Saint-Laurent, sont particulièrement menacées par une exposition aux contaminants car de grands navires transportant du titane et du fer passent dans l'archipel pour se rendre à Havre-Saint-Pierre durant toute l'année (Y. Aubry, comm. pers., 2015). À la suite d'un déversement causé par un navire, documenté en mars 1999, la nappe de pétrole ainsi produite a atteint la rive de la région de Mingan (Niles et al., 2008). L'exploration pétrolière et gazière s'est intensifiée le long du littoral nord-est et nord du Brésil (Paschoa, 2013), tandis que l'exploration pétrolière est en cours au Suriname et en Guyane (Morrison et al., 2012). De grandes installations pétrolières, comportant des puits à terre ou en mer, sont présentes près de lieux d'hivernage importants du rufa dans les secteurs chilien et argentin de la Terre de Feu et peuvent être à l'origine de catastrophes (R.I.G. Morrison et R.K. Ross, données inédites). Deux déversements d'hydrocarbures par des navires pétroliers ont été signalés près de l'entrée du détroit de Magellan (Niles et al., 2005), et de petites quantités d'hydrocarbures ont été observées sur des bécasseaux capturés durant des travaux de baguage dans la baie Lomas (A. Dey et L.J. Niles, données inédites). L'exploration pétrolière et l'exploitation du minerai de fer et de l'or, qui peuvent entraîner une pollution par les hydrocarbures et le mercure ainsi qu'une perte d'habitat, sont d'importantes menaces dans la partie centrale de la côte nord du Brésil et pourraient toucher la population brésilienne du Maranhão du rufa (Niles et al., 2005). L'importante halte migratoire de San Antonio Oeste, en Argentine, est aussi exposée à un risque de pollution du fait de la présence d'une fabrique de carbonate de soude (qui peut libérer jusqu'à 250 000 tonnes ou plus de chlorure de calcium par année, ce qui peut toucher les populations d'invertébrés intertidaux dont s'alimentent les oiseaux) et d'activités portuaires (pollution liée au transport maritime).
9.3 Effluents agricoles et sylvicoles (rufa et roselaari) :
Au Canada, un petit nombre de Bécasseaux maubèches pourrait être exposé à des herbicides et à des pesticides provenant d'activités agricoles en amont de la baie de Fundy (WHSRN, 2015). Les Bécasseaux maubèches, et leurs proies, peuvent être exposés à des effluents agricoles toxiques associés à la gestion des rizières à Trinidad, en Uruguay, en Argentine et en Guyane française (Blanco et al., 2006; Niles, 2012b; U.S. Fish and Wildlife Service, 2014). Les individus hivernant à l'embouchure du fleuve Colorado peuvent être particulièrement dérangés par les effluents agricoles provenant des États-Unis et du Mexique (G. Donaldson, comm. pers., 2015).
9.4 Déchets solides et ordures (rufa) :
Une décharge de la ville en croissance de Río Gallegos, en Argentine, se trouve à côté d'importantes aires de repos et d'alimentation du rufa (Ferrari et al., 2002). Les forts vents déposent les déchets dans de grandes portions de l'estuaire et cela diminue la qualité de l'habitat du Bécasseau maubèche (Ferrari et al., 2002). L'élimination non gérée de déchets solides dans la ville de Río Grande, en Argentine, menace l'habitat d'hivernage du rufa sur la Costa Atlantica (Rare, 2010).
11. Changements climatiques et phénomènes météorologiques violents
11.1 Déplacement et altération de l'habitat (rufa, roselaari et islandica) :
L'effet des changements climatiques sur le Bécasseau maubèche et sur ses proies est difficile à prévoir. Ces changements pourraient avoir divers effets positifs, neutres ou négatifs. Ces derniers pourraient quant à eux évoluer au fil du temps et/ou selon l'étendue des changements environnementaux. L'Arctique s'est réchauffé plus que toute autre région ces 30 dernières années (NSID, 2015); il fait donc partie des régions les plus susceptibles d'être touchées par les changements climatiques (ACIA, 2004). Meltofte et al. (2007) ont examiné en profondeur les effets possibles des changements climatiques sur les oiseaux de rivage dans l'Arctique. Parmi les principales préoccupations, on compte des changements dans l’habitat, (c’est-à-dire une contraction à long terme des milieux de l’Extrême-Arctique) et la désynchronisation de la phénologie des ressources alimentaires et des périodes de reproduction (c’est-à-dire que la disponibilité des ressources alimentaires ne coïncide pas avec la période de migration). En outre, les principales proies du Bécasseau maubèche pourraient être touchées de plusieurs autres façons. L'acidification de l'océan pourrait, par exemple, contribuer au déclin des proies qui dépendent largement du calcium, dont les mollusques bivalves (Byrne et Przeslawski, 2013, Parker et al., 2013), et les changements climatiques pourraient provoquer la hausse de l'activité des maladies en milieu marin, où des épidémies pourraient avoir des incidences négatives sur le Bécasseau maubèche et ses proies (Burge et al., 2014). Comme les limites de l'Extrême-Arctique devraient être refoulées vers le nord, les Bécasseaux maubèches, qui y nichent, seraient probablement parmi les espèces les plus touchées. Les populations se reproduisant dans la partie sud de l'Arctique, comme les rufa, qui nichent dans le centre de l'Arctique canadien, seraient les plus touchées. Les ruptures du cycle prédateurs-rongeurs, causées par les changements climatiques, peuvent entraîner des périodes prolongées de déprédation accrue sur le Bécasseau maubèche (c.-à-d. les adultes nicheurs, leurs oeufs et oisillons) (Meltofte et al., 2007; Niles et al., 2008; Fraser et al., 2013).
On a prévu des pertes potentielles de milieux intertidaux dues à l'élévation du niveau de la mer de 20 à 70 % au cours du prochain siècle dans cinq sites clés des États-Unis, dont la baie du Delaware (60 % de perte d'habitat; Galbraith et al., 2002). On prévoit une perte d'habitat à cause de l'élévation du niveau de la mer à la Terre de Feu (U.S. Fish and Wildlife Service, 2014), et d'autres sites seront probablement touchés. Les effets exacts sont difficiles à prévoir (IPCC, 2001; CCSP, 2009; U.S. Fish and Wildlife Service, 2014), mais des changements d'envergure aux berges sont prévus au cours des 100 prochaines années, ce qui met sérieusement en doute la capacité des sites de continuer à soutenir le nombre actuel d'oiseaux de rivage et fait ressortir la hausse des stress à venir sur les populations de bécasseaux.
Le rufa pourrait être avantagé par la hausse prévue des températures à court terme si les changements climatiques causent moins de retards dus au froid dans la fraie des limules de la baie du Delaware (Smith et Michels, 2006) et/ou la fonte prématurée de la neige recouvrant les lieux de reproduction de l'Arctique (Meltofte et al., 2007).
11.4 Tempêtes et inondations (rufa, roselaari et islandica) :
On observe une augmentation notable du nombre et de la force des ouragans partout dans le monde (1970–2004), notamment dans l'Atlantique Nord (Webster et al., 2005), dans les régions où le Bécasseau maubèche est présent (R.I.G. Morrison, données inédites). Des données de géolocalisation indiquent que les Bécasseaux maubèches modifient leur comportement de vol pour éviter certains systèmes météorologiques, ce qui augmente certainement leur dépense énergétique et peut influer sur leur survie (Niles et al., 2010). On ne sait pas si des bécasseaux ont effectivement été touchés (soit directement par la mortalité, soit indirectement par la diminution du nombre de proies dans les aires d'alimentation). Cependant, la hausse de la gravité des phénomènes météorologiques (incluant l'incidence accrue d'épisodes de précipitations abondantes [Fischer et Knutti, 2015]) présente certainement un risque accru, qui est susceptible d'augmenter d'après les prévisions de changements climatiques et la hausse de température des océans.
5. Objectifs en matière de population et de répartition (rufa et roselaari)/objectifs de gestion (islandica)
L’objectif à court terme en matière de population pour les sous-espèces rufa et islandica au Canada est de mettre fin au déclin de leur population d'ici 2025 à l'échelle du pays. L'objectif à long terme en matière de population pour le rufa sera ensuite d'augmenter, puis de maintenir la population à un niveau équivalent (ou supérieur) à ceux de 1986-1990 (100 000-150 000 individus [B. A. Harrington, résultats inédits dans Morrison et Harrington, 1992)]. L’objectif à long terme en matière de population pour l’islandica est de maintenir la population à son niveau actuel.
Les objectifs en matière de répartition des rufa et islandica nicheurs sont de maintenir la zone d'occurrence actuelle (c'est-à-dire la zone qui englobe l'aire de répartition géographique de toutes les populations nicheuses connues) au Canada. Un autre objectif en matière de répartition visant le rufa en migration est la conservation des haltes migratoires canadiennes abritant au moins 1 %Note4de bas de page de la population actuelle (1 % = 420 individus).
À la lumière des nouveaux renseignements concernant le roselaari obtenus depuis son évaluation par le COSEPAC en 2007 (c'est-à-dire on sait désormais que le roselaari qui se reproduit au Canada est en réalité le rufa, et seules quelques haltes migratoires mineures ont été trouvées au Canada), les objectifs en matière de population et de répartition sont de conserver le roselaari au Canada ainsi que toute halte migratoire canadienne abritant au moins 1 % de la population actuelle (1 % = 170 individus), ce qui favoriserait la persistance de l'espèce comme oiseau migrateur au Canada.
Les objectifs en matière de population portent sur le déclin à long terme de l'espèce, ce qui en a motivé la désignation (COSEWIC, 2007).
6. Stratégies et approches générales pour l'atteinte des objectifs
6.1 Orientation stratégique pour le rétablissement
Les activités de recherche et de conservation relatives au Bécasseau maubèche ont débuté dans les années 1970, et les travaux visant le rufa, en particulier, se sont intensifiés au milieu des années 1990 (pour en savoir plus, voir Dunan, 2014 : Manomet SRP/WHSRN, 2014). En 2009, un groupe de travail sur le Bécasseau maubèche a été mis sur pied pour les Amériques dans le cadre du Réseau de réserves pour les oiseaux de rivage dans l'hémisphère occidental (RRORHO). Le groupe s'est rassemblé pour cibler les besoins en matière de conservation et de recherche, pour établir des partenariats et des collaborations, ainsi que pour créer un plan visant le rétablissement du rufa (RRORHO, 2009).
L'orientation stratégique pour le rétablissement du rufa et du roselaari est décrite dans le tableau 3 tel qu'il est exigé pour les espèces en voie de disparition ou menacées faisant l'objet d'un programme de rétablissement. Le tableau 3 présente les approches de rétablissement et se fonde sur la vaste planification de la conservation déjà réalisée pour le Bécasseau maubèche à partir des plans suivants : le Plan canadien de conservation des oiseaux de rivage (Donaldson et al., 2000); les régions de conservation des oiseaux et les stratégies de conservation (ICOAN-Canada, 2015); la stratégie commerciale pour les oiseaux de rivage dans la voie migratoire de l'Atlantique (Atlantic Flyway Shorebird Business Strategy; Winn et al., 2013), document précurseur du plan d'affaires pour les oiseaux de rivage dans la voie migratoire de l'Atlantique (Atlantic Flyway Shorebird Business Plan), et le plan de conservation du Bécasseau maubèche pour l'hémisphère occidental (Red Knot Conservation Plan for the Western Hemisphere). D'autres renseignements détaillés et un calendrier de mise en œuvre suivront dans au moins un plan d'action.
Les mesures de conservation visant l'islandica sont détaillées dans le tableau 4 tel qu'il est exigé pour les espèces préoccupantes; le tableau contient aussi un calendrier de mise en œuvre de toutes les activités de conservation de la sous-espèce. Le tableau 4 s'inspire de la planification de la conservation déjà amorcée et des mesures de conservation proposées particulièrement pour l'islandica dans Leyrer et al., 2014.
| Menace ou élément limitatif | Stratégie générale pour le rétablissement | Prioriték | Description générale des approches de recherche et de gestion |
|---|---|---|---|
| Lacunes à combler dans les connaissances en vue du rétablissement | Suivi et recherche | Élevée |
|
| Toutes les menaces anthropiques | Conservation et gestion de l'espèce et de son habitat | Élevée |
|
| Toutes les menaces et les lacunes à combler dans les connaissances en vue du rétablissement | Éducation et sensibilisation, intendance et partenariats | Élevée |
|
| Toutes les menaces et les lacunes à combler dans les connaissances en vue du rétablissement | Éducation et sensibilisation, intendance et partenariats | Moyenne |
|
| Toutes les menaces anthropiques | Lois et politiques | Moyenne |
|
k « Priorité » reflète l'ampleur dans laquelle la stratégie générale contribue directement au rétablissement de l'espèce ou est un précurseur essentiel à une approche qui contribue au rétablissement de l'espèce.
| Stratégie générale | Mesure de conservation | Prioritél | Menaces ou préoccupations traitées | Échéance |
|---|---|---|---|---|
| Stratégie générale : conservation et gestion de l'espèce et de son habitat | Encourager l'élaboration de cadres fondés sur les voies migratoires et d'accords bilatéraux/multilatéraux qui encouragent les mesures coopératives de gestion et de protection des sites clés | Élevée | Toutes | Continue |
| Stratégie générale : conservation et gestion de l'espèce et de son habitat | Soutenir l'interdiction continue des pêches mécaniques dans la section néerlandaise de la mer des Wadden (dans le nord des Pays-Bas) | Faible | 5.4 Pêche et récolte de ressources aquatiques | Continue |
| Stratégie générale : conservation et gestion de l'espèce et de son habitat | Encourager les autorités responsables à interdire les pêches non durables qui ont des répercussions négatives sur l'espèce | Élevée | 5.4 Pêche et récolte de ressources aquatiques | Continue |
| Stratégie générale : conservation et gestion de l'espèce et de son habitat | Encourager les autorités responsables à atténuer les menaces que pose l'extraction pétrolière et gazière | Moyenne | 3.1 Forage pétrolier et gazier | Continue |
| Stratégie générale : éducation et sensibilisation, intendance et partenariats | Promouvoir la sensibilisation du public à l'espèce et aux menaces qui pèsent sur elle, notamment l'impact de la perturbation aux sites d'alimentation et de repos | Moyenne | Toutes les menaces anthropiques | Continue |
| Stratégie générale : lois et politiques | Promouvoir les mesures coopératives pour protéger l'espèce par des lois et promouvoir la conformité et/ou l'application des lois et règlements | Moyenne | 5.1 Chasse et capture d'animaux terrestres | Continue |
| Stratégie générale : suivi et recherche | Faciliter la recherche pour comprendre les menaces et les besoins en matière de conservation | Faible | Lacunes dans les connaissances en matière de rétablissement | Continue |
l « Priorité » reflète l'ampleur dans laquelle la stratégie générale contribue directement au rétablissement de l'espèce ou est un précurseur essentiel à une approche qui contribue au rétablissement de l'espèce.
6.2 Commentaires à l'appui du tableau de planification du rétablissement (rufa et roselaari), et des mesures de conservation et du calendrier de mise en œuvre (islandica)
Des renseignements contextuels supplémentaires sont fournis ci-dessous au sujet des approches de suivi et de recherche du tableau 3 qui n'atténuent pas précisément une menace. D'autres détails sur les approches de gestion figurent dans les nombreux documents qui existent sur la planification de la conservation du Bécasseau maubèche, mentionnés à la section 6.1. Ils seront présentés dans un ou plusieurs plans d'action subséquents pour le rufa et/ou le roselaari.
Le rétablissement d'une espèce qui possède une aire de répartition aussi étendue que le Bécasseau maubèche exigera l'engagement, la collaboration et la coopération à l'échelle nationale et internationale des autorités responsables fédérales, provinciales et territoriales, des conseils de gestion des ressources fauniques, des peuples autochtones, des collectivités locales, des propriétaires fonciers, de divers secteurs industriels et d'autres parties intéressées. Vu la dépendance du Bécasseau maubèche à l’égard de quelques sites clés ne servant pas à la reproduction, il sera important de suivre l’état de l’habitat, les tendances des populations et la répartition de l’espèce de sorte que l’efficacité des activités de rétablissement puisse être évaluée et améliorée, au besoin. Les programmes de suivi établis (p. ex. relevés aériens de la Terre de Feu) doivent être maintenus pour faire un suivi de l’état de certaines populations et l’efficacité des mesures de conservation.
Les activités de suivi et de recherche intensives du rufa sont menées dans l’ensemble de l’aire de répartition de cette sous-espèce ne servant pas à la reproduction depuis le milieu des années 1990 (Niles et al., 2007). En dépit de ces activités, on ne comprend pas bien les causes du déclin des populations. Les travaux sont pour la plupart non coordonnés, et il est nécessaire d'élaborer des protocoles et des plans de relevé normalisés pour étudier les caractéristiques des populations et de leur habitat. Des recherches visant à combler les nombreuses lacunes dans les connaissances doivent être réalisées pour assurer le rétablissement.
7. Habitat essentiel
L'habitat essentiel est l'habitat nécessaire à la survie ou au rétablissement d'une espèce. Aux termes de la LEP, la désignation et la protection de l'habitat essentiel s'appliquent uniquement aux espèces en voie de disparition et aux espèces menacées. Par conséquent, l'habitat essentiel n'est abordé dans le présent document que pour le rufa et le roselaari (c.-à-d. non pas pour l'islandica étant donné que son statut est celui d'espèce préoccupante). Ainsi, en vertu de l'alinéa 41(1)c) de la LEP, les programmes de rétablissement doivent inclure une désignation de l'habitat essentiel de l'espèce, dans la mesure du possible et selon la meilleure information accessible, ainsi que, des exemples d'activités susceptibles d'entraîner la destruction de l'habitat essentiel.
7.1 Désignation de l'habitat essentiel de l'espèce
L'habitat essentiel de reproduction du rufa ne peut être désigné pour le moment. L'utilisation de l'habitat et la répartition des aires de reproduction du rufa dans l'Arctique canadien sont mal définies parce que les nids des rufa sont cryptiques et difficiles à trouver, et que la répartition des rufa nicheurs est peu dense et couvre une superficie vaste et éloignée (U.S. Fish and Wildlife Service, 2014). Très peu de nids de la sous-espèce ont été trouvés au cours de décennies de recherche dans l'Arctique (J. Rausch et P. Smith, comm. pers., 2015), et il est impossible pour le moment d'effectuer des relevés à grande échelle pour cette espèce. La fidélité au site de nidification semble limitée pour cette sous-espèce (Niles et al., 2008); dans le cadre d'un projet de suivi sur 5 ans d'un petit site de l'île Southampton au Nunavut (Niles et al., 2008), le retour d'un seul mâle vers son territoire de reproduction a été documenté. Pour ces raisons, le degré d'incertitude entourant la désignation de l'habitat de reproduction nécessaire à la survie ou au rétablissement du rufa est élevé. Bien que des analyses préliminaires de la préférence en matière d'habitat aient été réalisées (Smith et Rausch, 2014), l'information accessible n'est pas adéquate pour permettre la désignation de l'habitat essentiel pour la reproduction (plus particulièrement, les données concernant les nids sont rares, et aucun examen au sol n'a été complété pour vérifier les hypothèses sur les préférences en matière d'habitat et éliminer les biais des travaux de recherche). L'habitat essentiel pour la reproduction ne s'applique pas au roselaari étant donné que rien n'indique que cette sous-espèce se reproduit au Canada.
D'après l'examen de l'aire de répartition géographique de l'espèce et de la spécificité de l'habitat, de l'abondance de la population et des menaces, l’habitat essentiel de halte migratoire du Bécasseau maubèche doit être désigné à l'échelle du site (c'est-à-dire aire de répartition géographique petite/localisée, spécificité limitée de l'habitat). Tout site utilisé par au moins 1 % de la population actuelle (c'est-à-dire rufa = 420 individus; roselaari = 170 individus) est désigné comme habitat essentiel de halte migratoire.
Les caractéristiques biophysiques connues de l’habitat essentiel de halte migratoire requises par le rufa sont les suivantes : milieux côtiers marins et estuariens vaseux, sablonneux ou rocheux présentant de vastes battures (p. ex. embouchure des baies et des estuaires, lagunes, marais salés, flèches de sable, îlots, hauts-fonds, bancs de sable, battures rocheuses [calcaires] couvertes ou non d'algues [p. ex. espèces du genre Fucus] et zones aux caractéristiques souvent associées aux inlets naturels) et/ou les milieux lacustres salés intérieurs. Les caractéristiques biophysiques de l'habitat de halte migratoire sont utilisées par les oiseaux à la fois, tant pour l'alimentation et le repos que le jour et la nuit. Pour l'alimentation, la sous-espèce doit avoir accès à une abondance de bivalves de taille adéquate et à d'autres invertébrés benthiques (c'est-à-dire organismes vivant dans les sédiments et/ou dans les couches sous la surface). Pour le repos, la sous-espèce a besoin d'avoir accès à un habitat, avec ou sans végétation, près des aires d'alimentation, présentant un espace adéquat pendant les marées les plus hautes et exempt de perturbations anthropiques excessives. Cet habitat doit aussi fournir une protection contre les prédateurs et le mauvais temps.
La désignation de l'habitat essentiel, dans le présent document, est considérée comme étant une désignation partielle et insuffisante pour répondre aux objectifs en matière de population et de répartition, parce qu'il est impossible d'établir avec certitude quel habitat est essentiel pour les oiseaux nicheurs, parce que les connaissances sur les haltes migratoires éloignées sont très limitées, et parce que l'importance de certains milieux utilisés comme haltes migratoires est actuellement inconnue (c'est-à-dire milieux dulcicoles des hautes terres). Un calendrier des études a été élaboré pour obtenir les renseignements nécessaires à l'achèvement de la désignation de l'habitat essentiel (voir la section 7.2).
Les zones renfermant l'habitat essentiel du rufa sont présentées dans les figures 2 à 14. L'habitat essentiel du rufa au Canada se trouve à l'intérieur des polygones ombrés en jaune (unités là où les critères et la méthodologie énoncés dans la présente section sont respectés). Le quadrillage UTM de 10 km sur 10 km montré dans ces figures est un système de quadrillage national de référence qui indique l'emplacement géographique général renfermant de l'habitat essentiel. Pour en savoir plus, communiquer avec Environnement et Changement climatique Canada – Service canadien de la faune au ec.planificationduretablissement-recoveryplanning.ec@canada.ca.
Aucun site ne répond actuellement aux critères de l'habitat essentiel de halte migratoire dans le cas du roselaari.

Description longue pour la figure 2
La figure 2 est une carte de l’habitat essentiel de halte migratoire du Bécasseau maubèche de la sous espèce rufa au lac Beaverhill, en Alberta. Il y a sur la carte six carrés du quadrillage UTM de référence de 10 kilomètres par 10 kilomètres, qui mettent en évidence l’emplacement géographique général de l’habitat essentiel de l’espèce. Les six carrés du quadrillage UTM renferment tous une portion de la Beaverhill Lake Heritage Rangeland Natural Area (zone naturelle des parcours patrimoniaux du lac Beaverhill), laquelle est la zone naturelle où se trouve l’habitat essentiel. Le parc national du Canada Elk Island est situé au nord ouest de la Beaverhill Lake Heritage Rangeland Natural Area.

Description longue pour la figure 3
La figure 3 est une carte de l’habitat essentiel de halte migratoire du Bécasseau maubèche de la sous espèce rufa au lac Reed, en Saskatchewan. Il y a sur la carte deux carrés du quadrillage UTM de référence de 10 kilomètres par 10 kilomètres, qui mettent en évidence l’emplacement géographique général de l’habitat essentiel de l’espèce. Les deux carrés du quadrillage UTM renferment chacun une portion du lac Reed, lequel est la zone naturelle où se trouve l’habitat essentiel. Le lac Reed est situé au sud ouest de Morse et au sud est de Herbert, deux municipalités de Saskatchewan.

Description longue pour la figure 4
La figure 4 est une carte de l’habitat essentiel de halte migratoire du Bécasseau maubèche de la sous espèce rufa au lac Chaplin, en Saskatchewan. Il y a sur la carte sept carrés du quadrillage UTM de référence de 10 kilomètres par 10 kilomètres, qui mettent en évidence l’emplacement géographique général de l’habitat essentiel de l’espèce. Les sept carrés du quadrillage UTM renferment chacun une portion du lac Chaplin, lequel est la zone naturelle où se trouve l’habitat essentiel. Le lac Chaplin est situé directement au sud de Chaplin, au sud ouest de Parkbeg et au sud est d’Ernfold, trois municipalités de Saskatchewan. Les Valjean Provincial Pastures (pâturages communautaires Valjean) et les Shamrock Community Pastures (pâturages communautaires Shamrock) sont des terres agricoles de la Couronne appartenant à la Province de Saskatchewan qui jouxtent le lac Chaplin.

Description longue pour la figure 5
La figure 5 est une carte de l’habitat essentiel de halte migratoire du Bécasseau maubèche de la sous espèce rufa aux lacs Quill, en Saskatchewan. Il y a sur la carte 15 carrés du quadrillage UTM de référence de 10 kilomètres par 10 kilomètres, qui mettent en évidence l’emplacement géographique général de l’habitat essentiel de l’espèce. Les 15 carrés du quadrillage UTM renferment chacun une portion d’un ou de plusieurs des lacs Quill, lesquels constituent la zone naturelle où se trouve l’habitat essentiel. Le Mount Hope-Prairie Rose Community Pasture (pâturage communautaire Mount Hope-Prairie Rose) est une terre agricole de la Couronne appartenant à la Province de Saskatchewan, qui borde l’habitat essentiel des lacs Quill et s’étend aussi au sud ouest de ces derniers. Les lacs Quill sont situés au nord de Wynyard, en Saskatchewan.

Description longue pour la figure 6
La figure 6 est une carte de l’habitat essentiel de halte migratoire du Bécasseau maubèche de la sous espèce rufa à Churchill, au Manitoba. Il y a sur la carte neuf carrés du quadrillage UTM de référence de 10 kilomètres par 10 kilomètres, qui mettent en évidence l’emplacement géographique général de l’habitat essentiel de l’espèce. Les neuf carrés du quadrillage UTM renferment chacun une portion de l’habitat essentiel, lequel s’étend sur la côte manitobaine de la baie d’Hudson entre Churchill et le parc national Wapusk. Le parc national Wapusk appartient au territoire domanial fédéral et se trouve à l’est de l’habitat essentiel représenté dans cette carte, et la baie d’Hudson se trouve immédiatement au nord de ce même habitat essentiel.

Description longue pour la figure 7
La figure 7 est une carte de l’habitat essentiel de halte migratoire du Bécasseau maubèche de la sous espèce rufa dans le parc national Wapusk, au Manitoba. Il y a sur la carte 11 carrés du quadrillage UTM de référence de 10 kilomètres par 10 kilomètres, qui mettent en évidence l’emplacement géographique général de l’habitat essentiel de l’espèce. Les 11 carrés du quadrillage UTM renferment chacun une portion de l’habitat essentiel, lequel s’étend sur la côte de la baie d’Hudson dans le parc national Wapusk, au sud est de Churchill. Tout l’habitat essentiel représenté dans cette carte se trouve dans le parc national Wapusk, qui appartient au territoire domanial fédéral.

Description longue pour la figure 8
La figure 8 est une carte de l’habitat essentiel de halte migratoire du Bécasseau maubèche de la sous espèce rufa dans le parc national Wapusk et au port Nelson, au Manitoba. Il y a sur la carte 19 carrés du quadrillage UTM de référence de 10 kilomètres par 10 kilomètres, qui mettent en évidence l’emplacement géographique général de l’habitat essentiel de l’espèce. Les 19 carrés du quadrillage UTM renferment chacun une portion de l’habitat essentiel, lequel s’étend sur la côte de la baie d’Hudson dans le parc national Wapusk, ainsi qu’au sud est du parc national Wapusk le long de la côte de la pointe Marsh. Neuf des carrés du quadrillage UTM renfermant de l’habitat essentiel se rapportent au parc national Wapusk, et les autres à la pointe Marsh.

Description longue pour la figure 9
La figure 9 est une carte de l’habitat essentiel de halte migratoire du Bécasseau maubèche de la sous espèce rufa dans le nord ouest de l’Ontario. Il y a sur la carte 13 carrés du quadrillage UTM de référence de 10 kilomètres par 10 kilomètres, qui mettent en évidence l’emplacement géographique général de l’habitat essentiel de l’espèce. Les 13 carrés du quadrillage UTM renferment chacun une portion de l’habitat essentiel, lequel s’étend sur la côte ontarienne de la baie d’Hudson entre la rivière Niskibi et la frontière entre l’Ontario et le Manitoba.

Description longue pour la figure 10
La figure 10 est une carte de l’habitat essentiel de halte migratoire du Bécasseau maubèche de la sous espèce rufa dans le parc provincial Polar Bear, en Ontario. Il y a sur la carte 13 carrés du quadrillage UTM de référence de 10 kilomètres par 10 kilomètres, qui mettent en évidence l’emplacement géographique général de l’habitat essentiel de l’espèce. Les 13 carrés du quadrillage UTM renferment chacun une portion de l’habitat essentiel, lequel s’étend sur la côte de la baie d’Hudson dans le parc provincial Polar Bear. Une certaine portion de chacun des carrés du quadrillage UTM est occupée par une partie du parc provincial.

Description longue pour la figure 11
La figure 11, qui est une vue d’ensemble des figures 11a à 11d, montre quatre zones distinctes d’habitat essentiel de halte migratoire du Bécasseau maubèche de la sous espèce rufa à la baie James.

Description longue pour la figure 11a
La figure 11a est une carte de l’habitat essentiel de halte migratoire du Bécasseau maubèche de la sous espèce rufa dans la portion québécoise de la baie James. Il y a sur la carte 10 carrés du quadrillage UTM de référence de 10 kilomètres par 10 kilomètres, qui mettent en évidence l’emplacement géographique général de l’habitat essentiel de l’espèce. Les 10 carrés du quadrillage UTM renferment chacun une portion de l’habitat essentiel, et le refuge d’oiseaux migrateurs de la Baie-Boatswain (aire protégée par le gouvernement fédéral) est englobé par 6 de ces 10 carrés du quadrillage UTM. Cette zone d’habitat essentiel est située au sud est de l’île Carey et de l’île Danby.

Description longue pour la figure 11b
La figure 11b est une carte de l’habitat essentiel de halte migratoire du Bécasseau maubèche de la sous espèce rufa dans la portion ontarienne de la baie James. Il y a sur la carte 28 carrés du quadrillage UTM de référence de 10 kilomètres par 10 kilomètres, qui mettent en évidence l’emplacement géographique général de l’habitat essentiel de l’espèce. Les 28 carrés du quadrillage UTM renferment chacun une portion de l’habitat essentiel, lequel s’étend sur la côte sud ouest de la baie James. Le refuge d’oiseaux migrateurs de la rivière Moose (aire protégée par le gouvernement fédéral) est englobé par les deux carrés du quadrillage UTM situés le plus au sud. Les 28 carrés du quadrillage UTM sont répartis du sud au nord depuis la rivière Moose jusqu’au-delà de Kashechewan.

Description longue pour la figure 11c
La figure 11c est une carte de l’habitat essentiel de halte migratoire du Bécasseau maubèche de la sous espèce rufa dans les portions ontarienne et nunavutoise de la baie James. Il y a sur la carte 41 carrés du quadrillage UTM de référence de 10 kilomètres par 10 kilomètres, qui mettent en évidence l’emplacement géographique général de l’habitat essentiel de l’espèce. Les 41 carrés du quadrillage UTM renferment chacun une portion de l’habitat essentiel, lequel s’étend sur la côte ouest de la baie James et le long des côtes ouest, sud et est de l’île Akimiski, au Nunavut. Le refuge d’oiseaux migrateurs de l’île Akimiski (aire protégée par le gouvernement fédéral) occupe les deux tiers de l’île Akimiski. Attawapiskat, en Ontario, se trouve à peu près au milieu, dans l’axe nord sud, de l’habitat essentiel représenté dans cette carte.
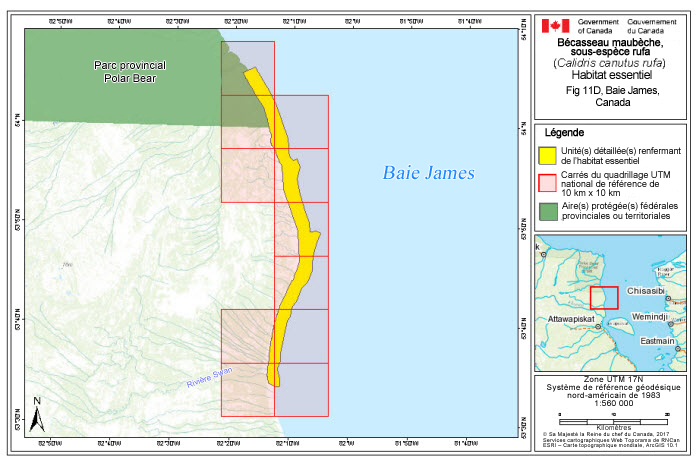
Description longue pour la figure 11d
La figure 11d est une carte de l’habitat essentiel de halte migratoire du Bécasseau maubèche de la sous espèce rufa dans la portion ontarienne de la baie James. Il y a sur la carte 11 carrés du quadrillage UTM de référence de 10 kilomètres par 10 kilomètres, qui mettent en évidence l’emplacement géographique général de l’habitat essentiel de l’espèce. Les 11 carrés du quadrillage UTM renferment chacun une portion de l’habitat essentiel, lequel s’étend sur la côte ouest de la baie James du sud au nord depuis environ la rivière Swan jusque dans le parc provincial Polar Bear.

Description longue pour la figure 12
La figure 12 est une carte de l’habitat essentiel de halte migratoire du Bécasseau maubèche de la sous espèce rufa le long du fleuve Saint Laurent, au Québec. Il y a sur la carte quatre carrés du quadrillage UTM de référence de 10 kilomètres par 10 kilomètres, qui mettent en évidence l’emplacement géographique général de l’habitat essentiel de l’espèce. Trois de ces carrés du quadrillage UTM renferment de l’habitat essentiel qui se trouve juste au sud du confluent de la rivière Saguenay avec le fleuve Saint Laurent, cet habitat essentiel se trouvant dans le parc marin du Saguenay–Saint-Laurent. L’autre carré du quadrillage UTM renferme de l’habitat essentiel qui se trouve sur une péninsule juste au sud de Portneuf-sur-Mer, au Québec.

Description longue pour la figure 13
La figure 13 est une carte de l’habitat essentiel de halte migratoire du Bécasseau maubèche de la sous espèce rufa dans l’archipel de Mingan, au Québec. Il y a sur la carte sept carrés du quadrillage UTM de référence de 10 kilomètres par 10 kilomètres, qui mettent en évidence l’emplacement géographique général de l’habitat essentiel de l’espèce. De l’habitat essentiel du Bécasseau maubèche se trouve à l’île Nue de Mingan, à quatre petites îles situées à l’ouest de cette dernière et, à l’est de celle ci, à la Grande Île et à l’île Quarry ainsi que sur une péninsule de la terre ferme située directement à l’ouest de la ville de Havre-Saint-Pierre. Tout l’habitat essentiel représenté dans cette carte se trouve dans la région de la côte nord du fleuve Saint Laurent, et, sauf pour ce qui est l’habitat essentiel se trouvant sur la péninsule de la terre ferme, à l’intérieur de la réserve de parc national de I’Archipel-de-Mingan, aire protégée par le gouvernement fédéral.

Description longue pour la figure 14
La figure 14 est une carte de l’habitat essentiel de halte migratoire du Bécasseau maubèche de la sous espèce rufa aux îles de la Madeleine, au Québec, entre le Nouveau-Brunswick et Terre-Neuve. Il y a sur la carte quatre carrés du quadrillage UTM de référence de 10 kilomètres par 10 kilomètres, qui mettent en évidence l’emplacement géographique général de l’habitat essentiel de l’espèce. L’une des deux zones d’habitat essentiel se trouve sur le littoral nord de l’île du Cap aux Meules et l‘autre sur le littoral sud ouest de cette même île.
7.2 Calendrier des études visant à désigner l'habitat essentiel
Un calendrier des études (tableau 5) a été élaboré afin de fournir l'information nécessaire à l'achèvement de la désignation de l'habitat essentiel nécessaire à l'atteinte des objectifs en matière de population et de répartition. La désignation de l'habitat essentiel sera mise à jour lorsque l'information deviendra accessible, soit dans un programme de rétablissement révisé, soit dans un plan d'action.
| Description de l'activité | Justification | Échéancier |
|---|---|---|
| 1. Habitat de reproduction : améliorer les connaissances sur l'utilisation de l'habitat par le rufa par la réalisation de relevés ciblés | Le Bécasseau maubèche se rencontre souvent dans des zones utilisées par quelques autres oiseaux de rivage et, par conséquent, peu de nids sont trouvés. Des relevés consacrés aux nids du Bécasseau maubèche fourniront une description des caractéristiques biophysiques de l'habitat nécessaires à sa reproduction, et pourraient renseigner sur la fidélité de l'espèce aux sites qu'elle utilise. | 2025 |
| 2. Habitat de reproduction : améliorer la modélisation de l'utilisation de l'habitat par le rufa | Les connaissances actuelles sur l'habitat essentiel sont fondées sur la classification grossière de l'habitat. Des données sur l'habitat à résolution spatiale améliorée sont accessibles, tout comme des techniques plus avancées de modélisation. L'application de ces données et méthodologie améliorées est en cours. Les données améliorées provenant des relevés ciblés amélioreront grandement les connaissances sur l'utilisation de l'habitat de reproduction. | 2017-2025 |
| 3. Habitat de reproduction : déterminer la limite septentrionale de l'aire de répartition de la sous-espèce rufa | L'islandica remplace le rufa vers le nord; il faut donc déterminer la limite septentrionale du rufa pour déterminer l'étendue de l'habitat essentiel de reproduction de la sous-espèce rufa. | En cours |
| 4. Habitat de halte migratoire : déterminer l'habitat de halte migratoire additionnel ainsi que son importance relative pour le Bécasseau maubèche (c'est-à-dire la proportion de chaque sous-population) au Canada | Les connaissances actuelles sur l'habitat de halte migratoire au Canada sont limitées par la difficulté d'accéder à des zones éloignées. De plus, c'est sur les haltes migratoires automnales que reposent les connaissances existantes. D'autres inventaires, relevés ainsi que l'utilisation de technologie en périodes de migration clés (printemps et automne) peuvent mener à la désignation d'habitat essentiel de halte migratoire additionnel. | 2025 |
7.3 Activités susceptibles d'entraîner la destruction de l'habitat essentiel
La destruction est déterminée au cas par cas. On peut parler de destruction lorsqu'il y a dégradation d'une partie de l'habitat essentiel, soit de façon permanente ou temporaire, à un point tel que l'habitat essentiel n'est plus en mesure d'assurer ses fonctions lorsque l'espèce en a besoin. La destruction peut découler d'une activité unique à un moment donné ou des effets cumulés d'une ou de plusieurs activités au fil du temps. Lorsque l'habitat essentiel est désigné dans un programme de rétablissement, des exemples d'activités susceptibles d'entraîner sa destruction sont fournis. Les exemples d'activités susceptibles d'entraîner la destruction de l'habitat essentiel de halte migratoire sont présentés dans le tableau 6.
| Description de l'activité | Description de l'effet |
|---|---|
| 1.1 Habitations et zones urbaines (p. ex. développement côtier ayant lieu dans un habitat de repos ou d'alimentation ou d'autres milieux étroitement liés à ces derniers); | Peut causer la destruction directe permanente ou temporaire de l'habitat (p. ex. la construction de ports et de quais, de chalets, de maisons ou d'hébergements touristiques, ou encore de promenades et de sentiers) et/ou avoir des effets indirects (p. ex. la modification du réseau hydrographique, l'altération de la salinité, des nutriments, de la topographie ou de l'oxygène dissous, etc. ce qui pourrait modifier la fonction des aires d'alimentation, et le compactage des sédiments qui pourrait avoir un impact sur les sources de nourriture). Les oiseaux pourraient être forcés à se déplacer vers des aires moins favorables ou non sécuritaires. |
| 1.2 Zones commerciales et industrielles (p. ex. construction de parcs éoliens ou de fermes houlomotrices dans un habitat de repos ou d'alimentation ou d'autres milieux étroitement liés à ces derniers); | Peut causer la destruction directe permanente ou temporaire de l'habitat et/ou avoir des effets indirects (p. ex. la modification du réseau hydrographique, l'altération de la salinité, des nutriments, de la topographie ou de l'oxygène dissous, etc. ce qui pourrait modifier la fonction des aires d'alimentation, et le compactage des sédiments qui pourrait avoir un impact sur les sources de nourriture). Les oiseaux pourraient être forcés à se déplacer vers des aires moins favorables ou non sécuritaires. |
| 2.4 Aquaculture en mer et en eau douce et 5.4 Pêche et récolte de ressources aquatiques (p. ex. récolte industrielle de bivalves); | Peut causer la destruction directe permanente ou temporaire de l'habitat et/ou avoir des effets indirects (p. ex. la perturbation et/ou la suppression des sources d'alimentation, la sédimentation qui pourrait avoir un impact sur la disponibilité de la nourriture). |
| 3.2 Exploitation de mines et de carrières (p. ex. procédés d'extraction et d'exploitation de sable); | Peut causer la destruction directe permanente de l'habitat d'alimentation et/ou de repos. |
| 6.1 Activités récréatives (p. ex. utilisation de véhicules hors route, tout terrain ou motorisés); | Peut causer la destruction directe permanente ou temporaire de l'habitat ou avoir des effets indirects (p. ex. la modification du réseau hydrographique, et le compactage des sédiments qui pourrait avoir un impact sur les sources de nourriture) forçant les oiseaux à se déplacer vers des aires d'alimentation ou de repos de faible qualité ou vers des aires non sécuritaires. |
| 7.2 Gestion et utilisation de l’eau et exploitation de barrages (p. ex. modification du régime d’écoulement) et 7.3 Autres modifications de l'écosystème (p. ex. remblayage de plage (c'est-à-dire l'ajout de sable à une berge érodée pour agrandir une plage existante, stabilisation d'une plage (structures rigides), nettoyage ou raclage des plages); | Peut causer la destruction directe permanente ou temporaire de l'habitat. Peut avoir des effets indirects permanents ou temporaires (p. ex. la perturbation et/ou la suppression d'une source de nourriture, la modification du réseau hydrographique, et le compactage des sédiments qui peut avoir un impact sur les sources de nourriture. Les oiseaux pourraient être forcés à se déplacer vers des aires moins favorables ou non sécuritaires. |
| 9.2 Effluents industriels et militaires (p. ex. déversement délibéré ou accidentel d'hydrocarbures, de pesticides ou de produits chimiques toxiques). | Peut causer la destruction permanente ou temporaire de l'habitat et/ou avoir des effets indirects (p. ex. la perturbation de sources de nourriture. |
8. Mesure des progrès
Les indicateurs de rendement présentés ci-dessous proposent un moyen de définir et de mesurer les progrès vers l'atteinte des objectifs en matière de population et de répartition.
- À court terme (c'est-à-dire avant 2025), les tendances à la baisse des populations pour le rufa et l'islandica ont cessé ou sont renversées.
- À long terme (c'est-à-dire après 2025), les populations de la sous-espèce rufa ont augmenté ou sont maintenues à des niveaux équivalents (ou supérieurs) à ceux de 1986-1990, et les populations de la sous-espèce islandica sont maintenues (soit aux niveaux actuels).
- La zone d'occurrence des rufa et des islandica est connue et maintenue au Canada, et celle du rufa est maintenue dans toutes les haltes migratoires canadiennes fréquentées par au moins 1 % de la population actuelle.
- À long terme (c’est-à-dire après 2025), le roselaari persiste en tant que migrateur au Canada.
9. Énoncé sur les plans d'action
Un ou plusieurs plans d'action visant le rufa et le roselaari seront publiés dans le Registre des espèces en péril dans les cinq ans suivant la publication du présent document.
10. Références
Abraham, K.F., et L.M. McKinnon. 2011. Hudson Plains Ecozone+ evidence for key findings summary. Canadian biodiversity: ecosystem status and trends 2010, evidence for key findings summary report No. 2. Canadian Councils of Resource Ministers. Ottawa, ON. vi + 98 p. [consulté en juillet 2015].
ACIA (Arctic Climate Impact Assessment). 2004. Impacts of a warming Arctic. [consulté en avril 2015].
Alaska Shorebird Group. 2008. Alaska shorebird conservation plan. Version II. Alaska Shorebird Group, Anchorage, AK. [consulté en avril 2015].
Aldabe J., E. Arballo, D. Caballero-Sadi, S. Claramunt, J. Cravino et P.I. Rocca. 2013. Aves. Pp. 149-173, dans: A. Soutullo, C. Clavijo, et J.A. Martínez-Lanfranco (eds.). Especies prioritarias para la conservación en Uruguay. Vertebrados, moluscos continentales y plantas vasculares. snap/dinama/mvotma y dicyt/mec, Montevideo. 222 pp.
Alvarez-Berrios, N.L., et T.M. Aide. 2015. Global demand for gold is another threat for tropical forests. Environ. Res. Lett. 10 014006. [consulté en mars 2015].
Andres, B.A., P.A. Smith, R.I.G. Morrison, C.L. Gratto-Trevor, S.C. Brown, et C.A. Friis. 2012. Population estimates of North American shorebirds, 2012. Wader Study Group Bulletin, 119(3): 178–194.
Atkinson, P.W., A.J. Baker, R.M. Bevan, N.A. Clark, K.B. Cole, P.M. González, J. Newton, L.J. Niles, et R.A. Robinson. 2005. Unravelling the migration and moult strategies of a long-distance migrant using stable isotopes: Red Knot Calidris canutus movements in the Americas. Ibis, 147: 738–749.
Atlantic Climate Adaptation Solutions. 2011. Climate change and shoreline protection in Atlantic Canada. [consulté en juillet 2015].
Atlantic States Marine Fisheries Commission. 2015. Horseshoe Crab. [consulté en avril 2015].
Azpiroz, A.B., et A. Rodríguez-Ferraro. 2006. Banded Red Knots Calidris canutus sighted in Venezuela and Uruguay. Cotinga 25:82-83.
Azpiroz, A.B., M. Alfaro et S. Jiménez. 2012. Lista roja de las aves de Uruguay. Una evaluación del estado de conservación de la avifauna nacional con base en los criterios de la Unión Internacional para la Conservación de la Naturaleza. Dirección Nacional de Medio Ambiente, Montevideo. 81 pp.
Baker, A.J., P.M. González, T. Piersma, L.J. Niles, d.N. de Lima Serrano, P.W. Atkinson, N.A. Clark, C.D.T. Minton, M.K. Peck, G. Aarts et al. 2004. Rapid population decline in red knots: Fitness consequences of decreased refuelling rates and late arrival in Delaware Bay. Proceedings of the Royal Society Biological Sciences, Series B 271(1541): 875-882.
Baker, A., P. González, R.I.G. Morrison, et B.A. Harrington. 2013. Red Knot (Calidris canutus). In A. Poole, ed.. The birds of North America Online. Cornell Lab of Ornithology, Ithaca.
Belton, W.1984. Birds of Rio Grande do Sul, Brazil. Part 1. Rehidae through Furnariidae. Bulletin of the American Museum of Natural History 178: 369-636.
Benoit, R. 2004. Centrale de l'Eastmain-1-A et dérivation Rupert. Avifaune – Limicoles migrateurs des baies de Rupert et Boatswain. Préparé pour la Société d'énergie de la baie James. Québec, FORAMEC inc. 95 p. et annexes.
BirdLife International 2012. Calidris canutus. The IUCN Red List of Threatened Species. Version 2014.3. [consulté en janvier 2015].
Blackburn, M., C.A.S. Mazzacano, C. Fallon, et S.Hoffman Black. 2014. Oil in our oceans: a review of the impacts of oil spills on marine invertebrates. The Xerces Society for Invertebrate Conservation, Portland, OR. 152 pp.
Blanco, D.E., B. López-Lanús, R.A. Dias, A. Azpiroz et F. Rilla. 2006. Use of rice fields by migratory shorebirds in southern South America. Implications for conservation and management. Wetlands International, Buenos Aires, Argentina.
Bocher, P., G. Quaintenne, P. Delaporte, C. Goulevant, B. Deceuninck, et E. Caillot. 2012. Distribution, phenology and long term trend of Red Knots Calidris canutus in France. Wader Study Group Bulletin, 119(1).
Botton, M.L., R.E. Loveland, et T.R. Jacobsen. 1994. Site selection by migratory shorebirds in Delaware Bay, and its relationship to beach characteristics and abundance of horseshoe crab (Limulus polyphemus) eggs. The Auk, 111(3): 605–616.
Brazil Wind Power. The largest wind power event in Latin America! 2015. Disponible à l'adresse : [consulté en avril 2015].
Bricelj, V.M., A.G. Haubois, M.R. Sengco, R. Pierce, J. Culter, et D.M. Anderson. 2012. Trophic transfer of brevetoxins to the benthic macrofaunal community during a bloom of the harmful dinoflagellate Karenia brevis in Sarasota Bay, Florida. Harmful Algae, 16: 27–34.
Buchanan, J.B., L.J. Salzer, G.E. Hayes, G. Schirato, et G.J. Wiles. 2010. Red Knot Calidris canutus migration at Grays Harbor and Willapa Bay, Washington: spring 2009. Wader Study Group Bulletin, 117: 41–45.
Buchanan, J.B., L.J. Salzer, G.J. Wiles, K. Brady, S.M. Desimone, et W. Michaelis. 2011. An investigation of Red Knot Calidris canutus spring migration at Grays Harbor and Willapa Bay, Washington. Wader Study Group Bulletin, 118: 97–104.
Buehler, D.M. et T. Piersma. 2008. Travelling on a budget: predictions and ecological evidence for bottlenecks in the annual cycle of long-distance migrants. Phi Trans R Soc B: 247-266
Buehler, D.M., L. Bugoni, G.M. Dorrestein, P.M. González, J. Pereira, Jr., L. Proença, I. de L. Serrano, A.J. Baker, et T. Piersma. 2010. Local mortality events in migrating sandpipers (Calidris) at a staging site in southern Brazil. Wader Study Group Bulletin, 117(3): 150–156.
Burge, C.A., C.M. Eakin, C.S. Friedman, B. Froelich, P.K. Hershberger, E.E. Hofmann, L.E. Petes, K.C. Prager, E. Weil, B.L. Willis, S.E. Ford, et C.D. Harvell. 2014. Climate change influences on marine infectious diseases: implications for management and society. Annual Review of Marine Science, 6: 249–277.
Burger, J., M. Gochfeld, et L. Niles. 1995. Ecotourism and birds in coastal New Jersey: contrasting responses of birds, tourists and managers. Environmental Conservation, 22: 56–64.
Burger, J., C. Jeitner, K. Clark, et K.J. Niles. 2004. The effect of human activities on migrant shorebirds: Successful adaptive management. Environmental Conservation 31(4):283-288.
Butler, R.W., R.C. Ydenberg et D.B. Lank. 2003. Wader migration on the changing predator land scape. Wader Study Group Bulletin 100:130-133.
Byrne, M. et R. Przeslawski. 2013. Multistressor Impacts of Warming and Acidification of the Ocean on Marine Invertebrates’ Life Histories. Integrative and Comparative Biology 53:582–596.
Carlos, C.J., C.E. Fedrizzi, A.A. Campos, H. Matthews-Cascon, C.X. Barroso, S.G. Rabay, L.E.A. Bezerra, C.A.O. Meirelles, A.J. Meireles, et P.R.L. Thiers. 2010. Migratory shorebirds conservation and shrimp farming in NE Brazil: final report, agreement # BR-N11. Rapport non publié préparé pour le U.S. Fish and Wildlife Service.
Carmona, R., N. Arce, V. Ayala-Perez, A. Hernández-Alvarez, J.B. Buchanan, L.J. Salzer, P.S. Tomkovich, J.A. Johnson, R.E. Gill, Jr., B.J. McCaffery, J.E. Lyons, L.J. Niles, et D. Newstead. 2013. Red Knot Calidris canutus roselaari migration connectivity, abundance and non-breeding distribution along the Pacific coast of the Americas. Wader Study Group Bulletin, 120(3): 168–180.
Castro, G., et J. P. Myers. 1993. Shorebird predation on eggs of horseshoe crabs during spring stopover on Delaware Bay. Auk 110:927-930.
CCSP (U.S. Climate Change Science Program). 2009. Thresholds of climate change in ecosystems. U.S. Climate Change Science Program synthesis and assessment product 4.2. U.S. Geological Survey, Reston, VA.
Clark, K.E., L.J. Niles, et J. Burger. 1993. Abundance and distribution of migrant shorebirds in Delaware Bay. The Condor, 95: 694–705.
Clark, K.E., R.R. Porter, et J.D. Dowdell. 2009. The shorebird migration in Delaware Bay. New Jersey Birds, 35(4): 85–92.
CMS (Convention on Migratory Species). 2005. Proposals for amendment of Appendices I and II of the Convention. In UNEP/CMS/Conf. 8.16 Annex, 5 October 2005, pp. 45–52,. Convention on Migratory Species, Bonn, Germany. [Également disponible en français : CMS (Convention sur la conservation des espèces migratrices appartenant à la faune sauvage). 2005 Propositions pour des amendements à apporter aux Annexes I et II de la Convention. In PNUE/CMS/Conf. 8.16 Annexe, le 5 octobre 2005, p. 45-52. Convention sur la conservation des espèces migratrices appartenant à la faune sauvage, Bonn, Allemagne).]
Cohen, J.B., S.M. Karpanty, J.D. Fraser, B.D. Watts, et B.R. Truitt. 2009. Residence probability and population size of red knots during spring stopover in the mid-Atlantic region of the United States. Journal of Wildlife Management 73(6):939-945.
Cohen, J.B., B.D. Gerber, S.M. Karpanty, J.D. Fraser, et B.R. Truitt, 2011. Day and night foraging of Red Knots (Calidris canutus) during spring stopover in Virginia, USA. Waterbirds, 34: 352–356.
COSEWIC. 2007. COSEWIC assessment and status report on the Red Knot Calidris canutus in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. vii + 58 p. [Également disponible en français : COSEPAC. 2007. Évaluation et Rapport de situation du COSEPAC sur le Bécasseau maubèche (Calidris canutus) au Canada, Comité sur la situation des espèces en péril au Canada, Ottawa, viii + 69 p.]
Davidson, N.C., et P.I. Rothwell. 1993. Disturbance to waterfowl on estuaries: the conservation and coastal management implications of current knowledge. Wader Study Group Bulletin, 68: 97–105.
Davis. 2012. Saving the amazing Red Knot from extinction -- it's complicated. [consulté en juillet 2015].
Dey, A.D., L.J. Niles, H.P. Sitters, K. Kalasz, et R.I.G. Morrison. 2011. Update to the status of the Red Knot Calidris canutus in the Western Hemisphere, April 2011. Draft update to the Status of the Red Knot (Calidris canutus rufa) in the Western Hemisphere. Studies in Avian Biology 36. Cooper Ornithological Society, CA. 14 pp.
DFO. 2012. Technical review of Baffinland's Mary River Project draft environmental impact statement (EIS). DFO Can. Sci. Advis. Sec. Sci. Advis. Rep. 2011/065 / DFO. [Également disponible en français : MPO. 2012a. Examen technique de l'énoncé des incidences environnementales (EIE) concernant la proposition de projet Mary River de Baffinland. Secr. can. de consult. sci. du MPO, Avis sci. 2011/065.]
Donaldson, G.M., C. Hyslop, R.I.G. Morrison, H.L. Dickson, et I. Davidson. 2000. Canadian Shorebird Conservation Plan. ISBN: 0-662-29112-3, Cat.: CW69-15/5-2000E. publications.gc.ca/.../collection_2011/ec/CW69-15-5-2000-eng.pdf [consulté en janvier 2015].
Environment Yukon. 2014. Yukon Species At risk. [consulté en janvier 2015].
Executive Office of the President. 2013. The President's Climate Action Plan. The White House, Washington, DC.
Ferrari S., C.Y. Albrieu, et P Gandini. 2002. Importance of the Rio Gallegos estuary, Santa Cruz, Argentina, for migratory shorebirds. Wader Study Group Bulletin, 99: 35-40.
Fischer, E.M., et R. Knutti. 2015. Anthropogenic contribution to global occurrence of heavy-precipitation and high-temperature extremes. Nature Climate Change, 5: 560-564.
Fraser, J.D., S.M. Karpanty, J.B. Cohen, et B.R. Truitt. 2013. The Red Knot (Calidris canutus rufa) decline in the western hemisphere: is there a lemming connection? Canadian Journal of Zoology, 91: 13–16.
Galbraith, H., R. Jones, R.A. Park, J.S. Clough, S. Herrod-Julius, B. Harrington, et G. Page. 2002. Global climate change and sea level rise: Potential losses of intertidal habitat for shorebirds. Waterbirds 25:173-183.
Godfrey, W.E. 1986. The birds of Canada. Revised Edition. National Museum of Natural Sciences, Ottawa. 595 pp. [Également disponible en français : Godfrey, W.E. 1986. Les oiseaux du Canada. Édition révisée. Musée national des sciences naturelles, Ottawa. 650 p.]
Godfrey, W.E. 1992. Subspecies of the Red Knot Calidris canutus in the extreme north-western Canadian arctic islands. Wader Study Group Bulletin, Supplement 64: 24–25.
González, P.M. 2005. Georgraphic area summary argentina: Report for developing a red knot status assessment in the U.S. Unpublished report by Fundacion Inalafquen, Rio Negro, Argentina.
Government of Canada. 2009. Species at Act Risk Policies: Overarching Policy Framework [Draft]. Species at Risk Act Policy and Guidelines Series. Environment Canada, Ottawa. 38 p. (Également disponible en français : Gouvernement du Canada. 2009. Politiques de la Loi sur les espèces en péril : Cadre général de politiques [ÉBAUCHE]. Gouvernement du Canada, Ottawa. ii + 42 p.).
Haramis, G.M., M.A. Teece, et D.B. Carter. 2002. Use of stable isotopes to determine the relative importance of horseshoe crabs in the diet of long-distance migrant shorebirds in Delaware Bay. Unpublished Report. Delaware Coastal Management Programs, Dover, DE.
Haramis, G.M., W.A. Link, P.C. Osenton, D.B. Carter, R.G. Weber, N.A. Clark, M.A. Teece, et D.S. Mizrahi. 2007. Stable isotope and penfeeding trial studies confirm value of horseshoe crab eggs to spring migrant shorebirds in Delaware Bay. Journal of Avian Biology, 38: 367–376.
Harrington, B.A. 1996. The flight of the red knot: a natural history account of a small bird's annual migration from the Arctic Circle to the tip of South America and back. W. W. Norton & Company, New York.
Harrington, B.A. 2001. Red knot (Calidris canutus). In A. Poole, and F. Gill, eds. The birds of North America, No. 563. The Birds of North America, Inc., Philadelphia, PA.
Henkel, J.R., B.J. Sigel, et C.M. Taylor. 2012. Large-scale impacts of the Deepwater Horizon oil spill: can local disturbance affect distant ecosystems through migratory shorebirds? BioScience, 62(7): 676–685.
Hernandez, D. 2005. Foraging efficiency of migratory shorebirds relative to horseshoe crab egg availability. Mémoire de maîtrise. 163 p. Rutgers University, NJ..
Hope, T.M., et C.E. Short. 1944. Southward migration of adult shorebirds on the west coast of James Bay, Ontario. The Auk, 61(4): 572–576.
IPCC (Intergovernmental Panel on Climate Change). 2001. Climate change 2001: impacts, adaptation and vulnerability. Chapter 6. Coastal zones and marine ecosystems. IPCC Secretariat, World Meteorological Organization, Geneva, Switzerland. [consulté en avril 2015].
Jackson, N.L., Nordstrom, K.F., et Smith, D.R., 2010, Armoring of estuarine shorelines and implications for horseshoe crabs on developed shorelines in Delaware Bay. In Shipman, H., Dethier, M.N., Gelfenbaum, G., Fresh, K.L., et Dinicola, R.S., eds., 2010, Puget Sound Shorelines and the Impacts of Armoring--Proceedings of a State of the Science Workshop, May 2009: U.S. Geological Survey Scientific Investigations Report 2010-5254, pp. 195-202.
Johnston, V. H., C. L. Gratto-Trevor, et S. T. Pepper. 2000. Assessment of bird populations in the Rasmussen Lowlands, Nunavut. Canadian Wildlife Service Occasional Paper No. 101, 56 pp. Service canadien de la faune, Ottawa.
Karpanty, S., J. Fraser, J.B. Cohen, et B.R. Truitt. 2014. Red knot use of coastal Virginia as a migration stopover site: 2013 Annual Report. Virginia Polytechnic Institute and State University, Blacksburg, VA.
Lathrop, R.G., Jr., L. Niles, D. Merchant, T. Farrell, et C. Licitra. 2013. Mapping the critical horseshoe crab spawning habitats of Delaware Bay. Rutgers Center for Remote Sensing & Spatial Analysis, New Brunswick, NJ.
Leighton, F.A. 1991. The toxicity of petroleum oils to birds: In J. White et L. Frink, eds. The effects of oil on wildlife: research, rehabilitation, and general concerns, pp. 43–57. The Oil Symposium, 16–18 October 1990. The Sheridan Press, Hanover, PA.
Leyrer, J., N. van Nieuwenhove, N. Crockford, et S. Delany. 2014. Proposals for concerted and cooperative action: bird species for consideration by COP11. UNEP/CMS/COP11/Inf 44. Report to the Convention on Migratory Species. [consulté en avril 2015]
López-Lanús, B., P. Grilli, E. Coconier, A. Di Giacomo et R. Banchs. 2008. Categorización de las aves de la Argentina según su estado de conservación. Informe de Aves Argentinas /AOP y Secretaría de Ambiente y Desarrollo Sustentable. Buenos Aires, Argentina.
Manning, T.H. 1952. Birds of the west James Bay and southern Hudson Bay coasts. National Museum of Canada Bulletin, No. 125: 1–114.
Manning, T.H., E.O. Hohn, et A.H. Macpherson. 1956. The birds of Banks Island. National Museum of Canada Bulletin, No. 143: 1–144.
Manning, T.H., et A.H. Macpherson. 1961. A biological investigation of Prince of Wales Island, N.W.T. Transactions of the Royal Canadian Institute, 33: 116–239.
Manomet SRP/WHSRN & the rufa Red Knot [PDF, 66.2 Ko]. 2014 [consulté en juillet 2015]. [en anglais seulement]
McGowan,.C., J.E. Hines, J.D. Nichols, J.E. Lyons, D.R. Smith et al. 2011. Demographic consequences of migratory stopover: linking red knot survival to horseshoe crab spawning abundance. Ecosphere 2, art 69.
McRae, R.D. 1982. Birds of Presqu'ile Ontario. Ministère des Richesses naturelles de l'Ontario, Ottawa.
Meireles, A.J.A., A. Gorayeb, D.R.F. Silva, et G.S. Lima. 2013. Socio-environmental impacts of wind farms on the traditional communities of the western coast of Ceará, in the Brazilian Northeast. Journal of Coastal Research, Special Issue No. 65: 81–86. ISSN 0749-0208.
Meltofte, H., T. Piersma, H. Boyd, B. McCaffery, B. Ganter, V.V. Golovnyuk, K. Graham, C.L. Gratto-Trevor, R.I.G. Morrison, E. Nol, et al. 2007. Effects of climate variation on the breeding ecology of Arctic shorebirds. Meddelelser om Grønland, Bioscience 59. Danish Polar Center, Copenhagen. 48 pp.
Morrison, R.I.G., et B.A. Harrington. 1992. The migration system of the Red Knot Calidris canutus rufa in the New World. Wader Study Group Bulletin, 64(Suppl.): 71–84.
Morrison, R I.G., B.J. McCaffery, R.E. Gill, S.K. Skagen, S.L. Jones, G.W. Page, C.L. Gratto-Trevor, et B.A. Andres. 2006. Population estimates of North American shorebirds, 2006. Wilson Journal of Ornithology (submitted manuscript), 1–76.
Morrison, R.I.G. 2006. Body transformations, condition, and survival in red knots Calidris canutus travelling to breed at Alert, Ellesmere Island, Canada. Ardea 94: 607-618.
Morrison, R.I.G., B.J. McCaffery, R.E. Gill, S.K. Skagen, S.L. Jones, G.W. Page, C.L. Gratto-Trevor, et B.A. Andres. 2007. Population estimates of North American shorebirds, 2006. Wader Study Bulletin, 111: 1–10.
Morrison, R.I.G., N.C. Davidson, et J.R. Wilson. 2007. Survival of the fattest: body stores on migration and survival in red knots Calidris canutus islandica. J Avian Biol 38: 479-487.
Morrison, R.I.G., D.S. Mizrahi, R.K. Ross, O.H. Ottema, N. de Pracontal, et A. Narine 2012. Dramatic declines of semipalmated sandpipers on their major wintering areas in the Guianas, Northern South America. Waterbirds, 35(1): 120–134.
Myers, J.P. 1986. Sex and gluttony on Delaware Bay. NaturalHistory, 95: 68–77.
NABCI Canada. 2015. Bird Conservation Regions and Conservation Strategies. [consulté en juin 2015]. [en anglais seulement]
Nagy, S., S. Flink, et T. Langendoen. 2014. Waterbird trends 1988-2012: results of trend analyses of data from the International Waterbird Census in the African-Eurasian Flyway (PDF; 6.1Mb). Wetlands International, Ede, the Netherlands. [consulté en juillet 2015]. [en anglais seulement]
Najjar, R.G., H.A. Walker, P.J. Anderson, E.J. Barron, R.J. Bord, J.R. Gibson, V.S. Kennedy, C.G. Knight, J.P. Megonigal, R.E. O'Connor, et al. 2000. The potential impacts of climate change on the mid-Atlantic coastal region. Climate Research, 14: 219–233.
NatureServe. 2015. NatureServe explorer: an online encyclopedia of life [web application]. Version 5.0. NatureServe, Arlington, VA. [consulté en janvier 2015]. [en anglais seulement]
Niles, L., A. Dey, H. Sitters, et C. Minton. 2005. Report on the status of red knots on the Delaware Bay with recommendations for the 2005 field season. NJDEP, Division of Fish and Wildlife, Endangered and Nongame Species Program, Trenton, NJ.
Niles, L. J., H. P. Sitters, A. D. Dey, P. W. Atkinson, A. J. Baker, K. A. Bennett, K. E. Clark, N. A. Clark, C. Espoz, P. M. González, B. A. Harrington, D. E. Hernández, K. S. Kalasz, R. N. Matus, C. D. T. Minton, R. I. G. Morrison, M. K. Peck, et I. L. Serrano. 2007. Status of the Red Knot (Calidris canutus rufa) in the Western Hemisphere. U.S. Fish & Wildlife Service, Pleasantville, NJ.
Niles, L.J., H.P. Sitters, A.D. Dey, P.W. Atkinson, A.J. Baker, K.A. Bennett, R. Carmona, K.E. Clark, N.A. Clark, C. Espoz, P.M. González, B.A. Harrington, D.E. Hernández, K.S. Kalasz, R.G. Lathrop, R.N. Matus, C.D.T. Minton, R.I.G.
Morrison, M.K. Peck, W. Pitts, R.A. Robinson, et I.L. Serrano. 2008. Status of the Red Knot, Calidris canutus rufa, in the Western Hemisphere. Studies in Avian Biology, 36: 1–185.
Niles, L.J. 2012. A rube with a view: The challege of the rice fields of Mana. [consulté en janvier 2015]. [en anglais seulement]
Niles, L.J., J.A.M. Smith, D.F. Daly, T. Dillingham, W. Shadel, A.D. Dey, M.S. Danihel, S. Hafner, et D. Wheeler. 2013a. Restoration of horseshoe crab and migratory shorebird habitat on five Delaware Bay beaches damaged by Superstorm Sandy (PDF, 4 Mb). [en anglais seulement]
Niles, L., T. Dillingham, D. Daly, J. Smith, A. Dey, et S. Hafner. 2013b. DRAFT: Creating resilient beach and marsh on Delaware Bay for shorebirds and horseshoe crabs: seven restoration projects for the future.
NSID (National Snow & Ice Data Centre). 2015. Climate change in the Arctic. [consulté en avril 2015]. [en anglais seulement]
Ouellet, H. 1969. Les oiseaux de l'île Anticosti, province de Québec, Canada. Musées nationaux du Canada, Musée national des sciences naturelles, Ottawa. Publications en zoologie no. 1, 79 p.
Parker, L.M., P.M. Ross, W.A. O'Connor, H.O. Pörtner, E. Scanes, et J.M. Wright. 2013. Predicting the response of molluscs to the impact of ocean acidification. Biology, 2: 651–692.
Parmelee, D.F., H.A. Stephens, et R.H. Schmidt. 1967. The birds of southeastern Victoria Island and adjacent small islands. National Museum of Canada Bulletin, 222: 1-229.
Paschoa, C. 2013. North Brazil oil -- deepwater oil off the State of Pará. Marine Technology News. [consulté en avril 2015]. [en anglais seulement]
Peterson, C.H., S.D. Rice, J.W. Short, D. Esler, J.L. Bodkin, B.E. Ballachey, et D.B. Irons. 2003. Long-term ecosystem response to the Exxon Valdez oil spill. Science, 302: 2082–2086.
Peterson, C.H., M.J. Bishop, L.M. D'Anna, et G.A. Johnson. 2014. Multi-year persistence of beach habitat degradation from nourishment using coarse shelly sediments. Science of the Total Environment, 487: 481–492.
Piersma, T., R. Hoekstra, A. Dekinga, A. Koolhaas, P. Wolf, P. Battley, et P. Wiersma. 1993. Scale and intensity of intertidal habitat use by knots Calidris canutus in the western Wadden Sea in relation to food, friends and foes. Netherlands Journal of Sea Research 31(4):331-357.
Piersma, T., G.A. Gudmundsson, et K. Lilliendahl. 1999. Rapid changes in the size of different functional organ and muscle groups during refueling in a long-distance migrating shorebird. Physiological and Biochemical Zoology 72(4):405-415.
Pizzolato, L., S.E.L. Howell, C. Derksen, J. Dawson, et L. Copland. 2014. Changing sea ice conditions and marine transportation activity in Canadian Arctic waters between 1990 and 2012. Climatic Change, 123 (2): 161–173.
Pomeroy, A.C., R.W. Butler, et R.C. Ydenberg. 2006. Experimental evidence that migrants adjust usage at a stopover site to trade off food and danger. Behavioral Ecology, 17(6):1041-1045.
Pravettoni, R. (UNEP/GRID-Arendal). 2011. Global Flyways of the six subspecies of Red Knot. Living Planet: Connected Planet, Rapid
Response Assessment. Disponible à l'adresse : www.grida.no/graphicslib/detail/global-yways-of-the-six-subspecies-of-red-knot_6683 [consulté en mars 2015].
Rare. 2010. Protecting the winter habitat of the famed Red Knot. Program Brochure. Rare, Arlington, VA.
RECHARGE. 2015. Brazil minister signals official interest in offshore wind. [consulté en avril 2015].
Resource Development Council. 2015. Alaska's oil & gas industry. [consulté en janvier 2015].
Ross, R.K., K. Abraham, R. Clay, B. Collins, J. Iron, R. James, D. McLachlin et R. Weeber. 2003. Ontario shorebird conservation plan. Environment Canada, Downsview, ON, Canada. [Également disponible en français : Ross, R.K., K. Abraham, R. Clay, B. Collins, J. Iron, R. James, D. McLachlin et R. Weeber. 2003. Plan de conservation des oiseaux de rivage de l'Ontario. Environnement Canada, Downsview (Ontario), Canada.]
Salafsky, N., D. Salzer, A.J. Stattersfield, C. Hilton-Taylor, R. Neugarten, S.H.M. Butchart, B. Collen, N. Cox, L.L. Master, S. O'Connor, et D. Wilkie. 2008. A standard lexicon for biodiversity conservation: unified classifications of threats and actions. Conservation Biology, 22: 897–911.
Schlacher, T.A., R. Noriega, A. Jones, et T. Dye. 2012. The effects of beach nourishment on benthic invertebrates in eastern Australia: impacts and variable recovery. Science of the Total Environment, 435-436:411-417.
Scherer, A.L. et M.V. Petry. 2012. Seasonal variation in shorebird abundance in the state of Rio Grande do Sul, southern Brazil. Wilson Journal of Ornithology 124: 40-50.
Siok, D., et B. Wilson. 2011. Using dredge spoils to restore critical American horseshoe crab (Limulus polyphemus) spawning habitat at the Mispillion Inlet. Delaware Coastal Program, Dover, DE.
Sitters, H. 2001. Notes on sites where red knots fed at low water and roosted at high water in the Atlantic coast wetlands, near Stone Harbor, New Jersey, during May 2001. Unpublished report to the Endangered and Nongame Species Program, New Jersey Division of Fish and Wildlife.
Skagen, S.K., et G. Thompson. 2013. Northern Plains/Prairie Potholes Regional Shorebird Conservation Plan. U.S. Shorebird Conservation Plan, Lakewood, CO.
Sklar, F.H., et J.A. Browder. 1998. Coastal environmental impacts brought about by alterations to freshwater flow in the Gulf of Mexico. Environmental Management, 22: 547–562.
Smith, D.R., et S.F. Michels. 2006. Seeing the elephant: importance of spatial and temporal coverage in a large-scale volunteer-based program to monitor horseshoe crabs. Fisheries, 31(10): 485–491.
Smith, P., et J. Rausch, 2014. Notes on habitat for Red Knot. Rapport interne préparé pour Environnement Canada.
Smith, L.C., et S.R. Stephenson. 2013. New trans-Arctic shipping routes navigable by midcentury. PNAS, 110(13): 4871–4872. doi: 10.1073/pnas.1214212110.
Sorenson, L., et L. Douglas. 2013. She did not die in vain. [consulté en avril 2015].
Soto-Montoya, E., R. Carmona, M. Gómez, V. Ayala-Pérez, N. Arce, et G.D. Danemann. 2009. Over-summering and migrant red knots at Golfo de Santa Clara, Gulf of California, Mexico. Wader Study Group Bulletin, 116(3): 191–194.
Stillman, R.A., A.D. West, J.D. Goss-Custard, S. McGrorty, N.J. Frost, D.J. Morrisey, A.J. Kenny, et A.L. Drewitt. 2005. Predicting site quality for shorebird communities: a case study on the Humber Estuary, UK. Marine Ecology Progress Series, 305: 203-217.
Tsipoura, N., et J. Burger. 1999. Shorebird diet during spring migration stopover on Delaware Bay. Condor, 101/3: 635-644.
U.S. Fish and Wildlife Service. 2011. Endangered and Threatened wildlife and plants; 90-day finding on a petition to list the Red Knot subspecies Calidris canutus roselaari as Endangered. Federal Register, vol. 76, No. 2. [consulté en janvier 2015].
U.S. Fish and Wildlife Service. 2014. rufa Red Knot ecology and abundance. Supplement to Endangered and Threatened wildlife and plants; proposed Threatened status for the rufa Red Knot (Calidris canutus rufa). [consulté en janvier 2015].
van Gils, J.A., P.F. Battley, T. Piersma, et R. Drent. 2005a. Reinterpretation of gizzard sizes of red knots world-wide emphasis overriding importance of prey quality at migratory stopover sites. Proceedings of the Royal Society of London, Series B 272:2609-2618.
van Gils, J.A., A. Dekinga, B. Spaans, W.K. Vahl, et T. Piersma. 2005b. Digestive bottleneck affects foraging decisions in red knots (Calidris canutus). II. Patch choice and length of working day. Journal of Animal Ecology, 74: 120–130.
Watts, B. 2009. Conservation in Conflict: Peregrines vs. Red Knots. [consulté en août 2015].
Webster, P.J., G.J. Holland, J.A. Curry, et H.R. Chang. 2005. Changes in tropical cyclone number, duration, and intensity in a warming environment. Science 309: 1844-1846.
Weir, R.D. 1989. Birds of the Kingston Region. Kingston Field Naturalists and Quarry Press Inc., Kingston, ON.
West, A.D., J.D. Goss-Custard, R.A. Stillman, R.W.G. Caldow, S.E.A. le V. dit Durell, et S. McGrorty. 2002. Predicting the impacts of disturbance on shorebird mortality using a behaviour-based model. Biological Conservation, 106(3): 319–328.
Wetlands International. 2015. Estimations des populations d’oiseaux d’eau. [consulté en janvier 2015].
WHSRN (Western Hemisphere Shorebird Reserve Network). 2009. 1st Meeting of the Red Knot (Calidris canutus) Working Group. [consulté en janvier 2015].
WHSRN (Western Hemisphere Shorebird Reserve Network). 2015. Site profiles. [consulté en avril 2015].
Winn, B, S. Brown, C. Spiegel, D. Reynolds, S. Johnston, et al. 2013. The Atlantic Flyway Shorebird Business Strategy. Disponible à l'adresse : manometcenter.pairserver.com/sites/default/files/publications_and_tools/AtlanticFlywayShorebirdBusinessStrategy.pdf [consulté en janvier 2015].
Zimmerling, J. R., A. C. Pomeroy, M. V. d'Entremont, et C. M. Francis. 2013. Canadian estimate of bird mortality due to collisions and direct habitat loss associated with wind turbine developments. Avian Conservation and Ecology 8(2): 10. [consulté en janvier 2015].
Communications personnelles
Y. Aubry. 2015. Biologiste, Environnement Canada, Québec, QC.
V. Brownell, J. Fitzsimmons, C. Risley, A. Tamachi, A Wheeldon, R. Donley. 2015. Ministère des Richesses naturelles et des Forêts, ON.
G. Donaldson. 2015. Gestionnaire, conservation des populations, Environnement Canada, Sackville, NB.
A. Duncan. 2015. Ligue pour la Protection des Oiseaux (LPO), France.
P.M. González. 2015. South American Shorebird Coordinator, International Conservation Fund of Canada. Río Negro, Argentine.
C.L. Gratto-Trevor. 2015. Direction générale des sciences et de la technologie, Environnement Canada, Saskatoon, SK.
V. Johnston. 2005. Biologiste des oiseaux de rivage, Environnement Canada, Yellowknife, NT.
M. Lamont. 2015. Technicien de la faune, ministère de l'Environnement, gouvernement du Nunavut, Kugluktuk, NT.
P. Marra. 2015. Chef. Migratory Bird Centre, Smithsonian Conservation Biology Institute, Washington, DC.
R.I.G. Morrison. 2015. Scientifique émérite, Direction générale des sciences et de la technologie, Environnement Canada, Ottawa, ON.
J. Rausch. 2015. Biologiste des oiseaux de rivage, Environnement Canada, Yellowknife, NT.
P. Smith. 2015. Direction générale des sciences et de la technologie, Environnement Canada, Ottawa, ON.
Annexe A: Effets sur l'environnement et sur les espèces non ciblées
Une évaluation environnementale stratégique (EES) est effectuée pour tous les documents de planification du rétablissement en vertu de la LEP, conformément à la Directive du Cabinet sur l'évaluation environnementale des projets de politiques, de plans et de programmes. L'objet de l'EES est d'incorporer les considérations environnementales à l'élaboration des projets de politiques, de plans et de programmes publics pour appuyer une prise de décisions éclairée du point de vue de l'environnement et d'évaluer si les résultats d'un document de planification du rétablissement peuvent affecter un élément de l'environnement ou tout objectif ou cible de la Stratégie fédérale de développement durable (SFDD).
La planification du rétablissement vise à favoriser les espèces en péril et la biodiversité en général. Il est cependant reconnu que des documents de planification du rétablissement peuvent, par inadvertance, produire des effets environnementaux qui dépassent les avantages prévus. Le processus de planification fondé sur des lignes directrices nationales tient directement compte de tous les effets environnementaux, notamment des incidences possibles sur des espèces ou des habitats non ciblés. Les résultats de l’EES sont directement inclus dans le document lui-même, mais également résumés dans le présent énoncé, ci-dessous.
Tous les oiseaux de rivage, par exemple le Bécasseau variable (Calidris alpina), le Tournepierre à collier (Arenaria interpres), le Bécasseau sanderling (Calidris alba), le Bécasseau semipalmé (Calidris pusilla), le Bécasseau roux (Limnodromus griseus) et le Pluvier siffleur (Charadrius melodus), qui dépendent des milieux côtiers marins et estuariens pour se nourrir et se reposer peuvent profiter de certaines des approches et/ou des mesures de conservation recommandées pour le Bécasseau maubèche. Les efforts visant à améliorer et/ou à restaurer l'habitat aux caractéristiques côtières fragiles pourraient être bénéfiques aux oiseaux de rivage migrateurs si de telles approches étaient jugées nécessaires et réalisables. Les mesures de rétablissement de l'espèce doivent être intégrées aux pratiques de gestion bénéfiques visant d'autres espèces inscrites, notamment là où de telles pratiques peuvent entrer en conflit.
La possibilité que les recommandations contenues dans le présent document entraînent des effets négatifs imprévus sur l'environnement et sur d'autres espèces a été examinée. La lutte contre les prédateurs risque de nuire à certaines espèces d'oiseaux de proie et de laridés, si une telle lutte est jugée réalisable et justifiée. Il a été conclu que les recommandations contenues dans le présent document n'entraîneront pas d'effets néfastes importants.
Annexe B: Besoins en recherche
- Améliorer les connaissances sur l'utilisation de l'habitat au moyen de relevés ciblés dans les lieux de reproduction (point no 1 du calendrier des études visant à désigner l'habitat essentiel).
- Améliorer les connaissances sur l'utilisation de l'habitat au moyen de relevés ciblés dans les aires d'alimentation et de repos utilisées par les bécasseaux, et déterminer le nombre d'individus utilisant les haltes dans différents sites au Canada (point no 4 du calendrier des études visant à désigner l'habitat essentiel).
- Améliorer les connaissances sur l'utilisation de l'habitat et les haltes migratoires des juvéniles (individus non-reproducteurs de moins de 2-3 ans) au moyen de relevés ciblés visant les concentrations après l'envol dans l'Arctique et à d’autres latitudes septentrionales, de même qu’à des latitudes méridionales (sud) et tropicales dans le cas des oiseaux dans leur première ou leur deuxième année (individus qui estivent potentiellement dans les aires d’hivernage).
- Recourir à la génétique, aux isotopes stables ou à d’autres techniques pour déterminer la sous-espèce des individus qui se trouvent dans la zone de chevauchement des lieux de reproduction de l’islandica/rufa dans l’Arctique canadien, et pour mieux délimiter l’habitat de reproduction de chacune de ces deux sous-espèces.
- Évaluer le statut actuel des populations ainsi que l’efficacité des mesures de conservation en effectuant des dénombrements annuels réguliers des populations dans les principaux lieux ne servant pas à la reproduction (p. ex. Terre de Feu, , Maranhao/Ceara, au Brésil, Guyane française, sud-est des États-Unis) et haltes migratoires (p. ex. péninsule Valdéz, San Antonio Oeste, Lagoa do Peixe, côte nord de l'Amérique du Sud, baie du Delaware, îles-barrières de la côte de la Virginie, archipel de Mingan, littoraux ouest et est de la baie James).
- Examiner la possibilité d'améliorer le suivi de la migration du Bécasseau maubèche au Canada en vue de compléter les données obtenues à partir du suivi « hivernal » en repérant toutes les haltes migratoires existantes dans chaque région, en tenant compte des éléments du plan de relevé (p. ex. sélection du site, optimisation des protocoles d'échantillonnage, variabilité annuelle de la qualité des haltes migratoires), en déterminant de manière périodique la durée du séjour et les facteurs de causalité associés dans des haltes migratoires données et, enfin, en évaluant les taux de détection pour réduire les biais d'échantillonnage.
- Poursuivre les travaux de marquage-recapture (observation d'individus avec marqueurs de type drapeaux codés) pour déterminer les variations du taux de survie annuelle, le stade du cycle vital marqué par le plus haut taux de mortalité (et pourquoi il en est ainsi) et l'efficacité des mesures de gestion, de même que pour comprendre les connexions entre l'habitat de reproduction, les haltes migratoires et l’habitat utilisé ne servant pas à la reproduction.
- Déterminer les causes du déclin de certaines populations dans certains sites en évaluant les effets de paramètres environnementaux et autres (p. ex. changements climatiques via les températures/tempêtes dans l'Arctique, période d'éclosion des insectes et des poussins, fréquence/survenue des ouragans pendant la migration, sécheresses/inondations), et évaluer les effets des prédateurs, des perturbations anthropiques, de la pression exercée par la chasse, des espèces problématiques (p. ex. surabondance des oies des neiges (Chen caerulescens) pendant la migration), des contaminants et de la modification de l'habitat en tant que sources des déclins observés.
- Examiner différents types de disponibilité de la nourriture et de méthodes de quête de nourriture dans les haltes migratoires clés le long de la côte de l'Atlantique et ailleurs pour préciser l'importance de la baie du Delaware (et du limule, espèce proie) par rapport à d'autres sites, et fournir de l'information sur la souplesse potentielle des modes de quête de nourriture du rufa (ou l'absence de souplesse).
- Examiner l'écologie de la reproduction, le comportement et la survie au nid des bécasseaux dans les lieux de reproduction arctiques pour déterminer si les conditions pendant la saison de reproduction (p. ex. conditions météorologiques et abondance des rongeurs microtinés) limitent les populations et comment, le cas échéant, les changements démographiques réagissent le plus aux variations du taux de survie des adultes.
- Recourir à la génétique, aux isotopes stables et à d'autres techniques pour déterminer le lieu de reproduction d'origine des individus.
Détails de la page
- Date de modification :