
Magdalen Islands Grasshopper (Melanoplus madeleineae): COSEWIC assessment and status report 2016
Official title: COSEWIC assessment and status report on the Magdalen Islands Grasshopper Melanoplus madeleineae in Canada

Special concern
2016
Table of contents
- Table of contents
- Assessment summary
- Executive summary
- Technical summary
- Preface
- Wildlife species description and significance
- Distribution
- Habitat
- Biology
- Population sizes and trends
- Threats and limiting factors
- Protection, status and ranks
- Acknowledgements
- Authorities contacted
- Information sources
- Biographical summary of report writers
- Collections examined
List of figures
- Figure 1. Female Magdalen Islands Grasshopper. Specimen collected from Île du Havre aux Maisons in the Magdalen Islands, Québec August 11, 2010. Specimen collected by Paul Catling and deposited at the Canadian National Collection of Insects, Arachnids and Nematodes, Ottawa, Ontario. Photo Cory Sheffield.
- Figure 2. Male Magdalen Islands Grasshopper. Specimen collected from Île du Havre aux Maisons in the Magdalen Islands, Québec August 11, 2010. Specimen collected by Paul Catling and deposited at the Canadian National Collection of Insects, Arachnids and Nematodes, Ottawa, Ontario. Photo Cory Sheffield.
- Figure 3. Location of Magdalen Islands in Canada and North America.
- Figure 4. The Magdalen Islands Grasshopper is confirmed most recently from Île du Havre aux Maisons and Île d’Entrée. Map GoogleEarth.
- Figure 5. Maximum extent of occurrence (EOO) and index of area of occupancy (IAO) calculations for the Magdalen Islands Grasshopper. Note, the grasshopper has not been recorded in all habitats on the islands. The spatial area of the islands is 205.5 km2.
- Figure 6. Search effort for the Magdalen Islands Grasshopper in 2015.
- Figure 7. Open grassy maritime meadow Magdalen Islands Grasshopper habitat. The flowering plants include Oxeye Daisy (Leucanthemum vulgare). L'échouerie de la Pointe Basse Cap Alrigh. Specimens were collected from this site in 2010 by Catling et al. (2013). Photo taken July 10, 2015 by Jennifer Heron.
- Figure 8. Open grassy maritime meadow Magdalen Islands Grasshopper habitat. L'échouerie de la Pointe Basse Cap Alright. Specimens were collected from this site in 2010 by Catling et al. (2013). Photo taken July 10, 2015 by Jennifer Heron.
- Figure 9. Open grassy maritime meadow where immature Magdalen Islands Grasshopper specimens were collected and barcoded, Île du Havre aux Maisons. Photo taken July 9, 2015 by Jennifer Heron.
- Figure 10. Open grassy maritime meadow where immature Magdalen Islands Grasshopper specimens were collected and barcoded, Chemin de la Belle-Anse off Chemin des Caps, Île du Cap aux Meules, Magdalen Islands. Photo taken July 10, 2015 by Jennifer Heron.
- Figure 11. Open grassy maritime meadow where immature Magdalen Islands Grasshopper specimens were collected and barcoded, Chemin de la Belle Anse off Chemin des Caps. Photo taken July 10, 2015 by Jennifer Heron.
- Figure 12. Map of the Magdalen Islands showing two sectors of habitat projected to be breached because of rising sea levels as a result of climate change. Map from Bernatchez et al. (2008).
List of tables
- Table 1. Magdalen Islands Grasshopper museum and collection records. [Editoral note: This table has been modified to remove geographic coordinates. The complete table can be obtained by contacting the COSEWIC Secretariat.]
- Table 2. Search effort for Magdalen Islands Grasshopper, July 6 – 12, 2015 totalled 40 person-hours over 24 sites. [Editoral note: This table has been modified to remove geographic coordinates. The complete table can be obtained by contacting the COSEWIC Secretariat.]
- Table 3. The International Union for Conservation of Nature – Conservation Measures Partnership (IUCN-CMP) threats calculator outputs for the Magdalen Islands Grasshopper.
Document information
COSEWIC
Committee on the Status
of Endangered Wildlife
in Canada
![]()
COSEPAC
Comité sur la situation
des espèces en péril
au Canada
COSEWIC status reports are working documents used in assigning the status of wildlife species suspected of being at risk. This report may be cited as follows:
COSEWIC. 2016. COSEWIC assessment and status report on the Magdalen Islands Grasshopper Melanoplus madeleineae in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. ix + 44 pp. (Species at Risk Public Registry website).
Production note:
COSEWIC would like to acknowledge Cory S. Sheffield and Jennifer Heron for writing the status report on the Magdalen Islands Grasshopper, Melanoplus madeleineae, in Canada, prepared under contract with Environment and Climate Change Canada. This report was overseen and edited by Paul Grant, Co-chair of the COSEWIC Arthropods Specialist Subcommittee.
For additional copies contact:
COSEWIC Secretariat
c/o Canadian Wildlife Service
Environment and Climate Change Canada
Ottawa, ON
K1A 0H3
Tel.: 819-938-4125
Fax: 819-938-3984
E-mail: COSEWIC E-mail
Website: COSEWIC
Également disponible en français sous le titre Ếvaluation et Rapport de situation du COSEPAC sur le Criquet des Îles-de-la-Madeleine (Melanoplus madeleineae) au Canada.
Cover illustration/photo:
Cover photograph by Paul Catling: Female Magdalen Islands Grasshopper photographed within open meadow habitat on Île du Havre aux Maisons in the Magdalen Islands, QC, with Île d’Entrée in the background on August 11, 2010.
COSEWIC Assessment summary
Assessment summary – November 2016
- Common name
- Magdalen Islands Grasshopper
- Scientific name
- Melanoplus madeleineae
- Status
- Special Concern
- Reason for designation
- This Canadian endemic is restricted to the Magdalen Islands in Quebec, where it is known to occur on seven of the eight main islands. Threats to this species are low, but recreational activities, road mortality and habitat loss through predicted coastal erosion may impact this species or its habitat.
- Occurrence
- Quebec
- Status history
- Designated Special Concern in November 2016.
COSEWIC Executive summary
Magdalen Islands Grasshopper
Melanoplus madeleineae
Wildlife species description and significance
Magdalen Islands Grasshopper (Melanoplus madeleineae) is a large (21 – 29 mm) nondescript species endemic to the Magdalen Islands, Québec, Canada. The main diagnostic features are on the hind femur, which is dark crimson on the lower surface, and uniformly dark coloured (i.e., non-banded) on the outer surface.
The Magdalen Islands Grasshopper is a relic of the Wisconsinan ice age. The species may have derived from the smaller, yet morphologically similar and closely related Northern Spur-throat Grasshopper, which does not occur on the Magdalen Islands yet is widespread on the mainland.
Distribution
The global and Canadian range of the Magdalen Islands Grasshopper is restricted to the Magdalen Islands within the Gulf of Saint Lawrence, Québec. The Magdalen Islands archipelago is composed of eight main islands and several other small uninhabited islands. This grasshopper has been recorded from seven of the main islands. Six of these (Île du Havre Aubert, Île d’Entrée, Île du Cap aux Meules, Grosse Île, Île du Havre aux Maisons and Pointe aux Loups) are connected by sand bars and smaller islands, whereas Île d’Entrée is separated by about 4 km from the main group. This species is not known from Brion Island, which is approximately 13 km from Grosse Île.
Habitat
The Magdalen Islands Grasshopper occurs within open maritime meadow and grass/sedge hillside habitats containing plant communities.
Biology
The Magdalen Islands Grasshopper overwinters as an egg, hatching as a nymph in the early spring. The species may have a two-year diapause, as has been reported with its sister taxon, the Northern Spur-throat Grasshopper. Growth is by gradual metamorphosis, with each of the five nymphal instars getting progressively larger, and with more pronounced morphological adult characteristics, as they moult. Both nymphs and adults share feeding habits. Adults are active from mid-July through to late September and mating and oviposition occur during this time. Females oviposit egg pods within soft soil substrates, on potentially bare ground such as trails and dirt roads. Like most spur-throated grasshoppers, this species probably feeds on a wide range of plant hosts, though specific feeding preferences (if any) are unknown.
Population size and trends
There is no information on subpopulation size(s) and trends for the Magdalen Islands Grasshopper. Approximately 80 specimens have been collected and deposited as museum vouchers. Surveys to date have focused on recording new subpopulations, collecting natural history and habitat information, and genetic studies. Strategic surveys for this species have been primarily by sweep-netting for grasshoppers and collecting in suitable habitats. It is difficult to time collection events to correspond with peak adult emergence, suggesting that detection success and perceptions of rarity could vary considerably.
Threats and limiting factors
There are several potential threats to the Magdalen Islands Grasshopper. Road mortality is potentially high for grasshoppers where roadways bisect suitable habitat or where adults may oviposit within the soft substrate of dirt roads. Recreational activities such as all-terrain vehicle operations on pathways through meadow habitats can cause direct mortality as well as compaction of soil and grasshopper forage plants. The species may also be subject to a range of natural predators, parasites and pathogens that regulate orthopteran populations. Overall threat impact is considered Low.
Protection, status, and ranks
There are no federal or provincial laws that specifically protect the Magdalen Islands Grasshopper, mitigate specific threats to grasshoppers, or protect this species’ habitat. The global conservation status rank is G2 (Imperilled), the Canadian national status rank is N2 (Imperilled) and the Québec provincial status is S2 (Imperilled). Approximately 30% of the main island group of the Magdalen Islands is public land, although the portion that is occupied by the Magdalen Islands Grasshopper is unknown.
Technical summary
- Scientific name:
- Melanoplus madeleineae
- English name:
- Magdalen Islands Grasshopper
- French name:
- Criquet des Îles-de-la-Madeleine
- Range of occurrence in Canada :
- Québec
Demographic information
| Summary items | Information |
|---|---|
| Generation time | 1 – 2 years. |
| Is there an [observed, inferred, or projected] continuing decline in number of mature individuals? | Unknown. |
| Estimated percent of continuing decline in total number of mature individuals within [5 years or 2 generations]. | Unknown. |
| [Observed, estimated, inferred, or suspected] percent [reduction or increase] in total number of mature individuals over the last [10 years, or 3 generations]. | Unknown. |
| Projected or suspected] percent [reduction or increase] in total number of mature individuals over the next [10 years, or 3 generations]. | Unknown. |
| [Observed, estimated, inferred, or suspected] percent [reduction or increase] in total number of mature individuals over any [10 years, or 3 generations] period, over a time period including both the past and the future. | Unknown. |
| Are the causes of the decline a) clearly reversible and b) understood and c) ceased? | a. Not applicable. b. Not applicable. c. Not applicable. |
| Are there extreme fluctuations in number of mature individuals? | Potentially, yes. |
Extent and occupancy information
| Summary items | information |
|---|---|
| Estimated extent of occurrence | 1021 km2 (based on occupancy over all islands where the species has been recorded). |
| Index of area of occupancy (IAO) (2 x 2 grid cells = 142). |
568 km2 (however, the land area of the islands is 205.5 km2) |
| Is the population “severely fragmented” i.e., is >50% of its total area of occupancy in habitat patches that are (a) smaller than would be required to support a viable population, and (b) separated from other habitat patches by a distance larger than the species can be expected to disperse? | a. No. b. Unknown. |
| Number of locations (Note: See Definitions and abbreviations on COSEWIC website and IUCN (Feb 2014) for more information on this term.) |
7 – 14 |
| Is there an [observed, inferred, or projected] decline in extent of occurrence? | No. |
| Is there an [observed, inferred, or projected] decline in index of area of occupancy? | No. |
| Is there an [observed, inferred, or projected] decline in number of subpopulations? | No. |
| Is there an [observed, inferred, or projected] decline in number of “locations”? (Note: See Definitions and abbreviations on COSEWIC website and IUCN (Feb 2014) for more information on this term.) |
No. |
| Is there an [observed, inferred, or projected] decline in [area, extent and/or quality] of habitat? | Yes. Inferred decline in habitat quality. |
| Are there extreme fluctuations in number of subpopulations? | No. |
| Are there extreme fluctuations in number of “locations”? (Note: See Definitions and abbreviations on COSEWIC website and IUCN (Feb 2014) for more information on this term.) |
No. |
| Are there extreme fluctuations in extent of occurrence? | No. |
| Are there extreme fluctuations in index of area of occupancy? | No. |
Number of mature individuals (in each subpopulation)
| Subpopulations (give plausible ranges) | N Mature Individuals |
|---|---|
| Total | Unknown. |
Quantitative analysis
| Summary items | Information |
|---|---|
| Probability of extinction in the wild is at least [20% within 20 years or 5 generations, or 10% within 100 years]. | Unknown. No data available to complete analysis. |
Threats
Was a threats calculator completed for this species? Yes. Overall low impact.
- 4.1 Roads and railroads (low impact)
- 6.1 Recreational activities (low impact)
- 8.1 Invasive non-native/alien species (low impact)
What additional limiting factors are relevant? Predation and parasitism.
Rescue effect (immigration from outside Canada)
| Summary items | Information |
|---|---|
| Status of outside population(s) most likely to provide immigrants to Canada. | Not applicable. The species is endemic to the Magdalen Islands. |
| Is immigration known or possible? | Not applicable. |
| Would immigrants be adapted to survive in Canada? | Not applicable. |
| Is there sufficient habitat for immigrants in Canada? | Not applicable. |
| Are conditions deteriorating in Canada? See Table 3 (Guidelines for modifying status assessment based on rescue effect). |
Not applicable. |
| Are conditions for the source population deteriorating? See Table 3 (Guidelines for modifying status assessment based on rescue effect). |
Not applicable. |
| Is the Canadian population considered to be a sink? See Table 3 (Guidelines for modifying status assessment based on rescue effect). |
Not applicable. |
| Is rescue from outside populations likely? | Not applicable. |
Data-sensitive species
| Summary items | Information |
|---|---|
| Is this a data sensitive species? | No. |
Status history
COSEWIC: Designated Special Concern in November 2016.
Status and reasons for designation:
| Summary items | Information |
|---|---|
| Status: | Special Concern |
| Alpha-numeric code: | Not applicable |
| Reasons for designation: | This Canadian endemic is restricted to the Magdalen Islands in Québec, where it is known to occur on seven of the eight main islands. Threats to this species are low, but recreational activities, road mortality and habitat loss through predicted coastal erosion may impact this species or its habitat. |
Applicability of criteria:
| Summary items | Information |
|---|---|
| Criterion A (Decline in total number of mature individuals): |
Not applicable. Insufficient data available. |
| Criterion B (Small distribution range and decline or fluctuation): |
This species is near to qualifying for Threatened based on restricted distribution, but could not meet the criteria with certainty. |
| Criterion C (Small and declining number of mature individuals): |
Not applicable. Insufficient data to estimate decline in total number of mature individuals. |
| Criterion D (Very small or restricted population): |
Not applicable. Insufficient data on number of mature individuals within population and the index of area of occupancy and number of locations are above the thresholds. |
| Criterion E (Quantitative analysis): |
Not applicable. Existing data insufficient to project population probability of extinction or extirpation in the wild. |
Preface
COSEWIC history
The Committee on the Status of Endangered Wildlife in Canada (COSEWIC) was created in 1977 as a result of a recommendation at the Federal-Provincial Wildlife Conference held in 1976. It arose from the need for a single, official, scientifically sound, national listing of wildlife species at risk. In 1978, COSEWIC designated its first species and produced its first list of Canadian species at risk. Species designated at meetings of the full committee are added to the list. On June 5, 2003, the Species at Risk Act (SARA) was proclaimed. SARA establishes COSEWIC as an advisory body ensuring that species will continue to be assessed under a rigorous and independent scientific process.
COSEWIC mandate
The Committee on the Status of Endangered Wildlife in Canada (COSEWIC) assesses the national status of wild species, subspecies, varieties, or other designatable units that are considered to be at risk in Canada. Designations are made on native species for the following taxonomic groups: mammals, birds, reptiles, amphibians, fishes, arthropods, molluscs, vascular plants, mosses, and lichens.
COSEWIC membership
COSEWIC comprises members from each provincial and territorial government wildlife agency, four federal entities (Canadian Wildlife Service, Parks Canada Agency, Department of Fisheries and Oceans, and the Federal Biodiversity Information Partnership, chaired by the Canadian Museum of Nature), three non-government science members and the co-chairs of the species specialist subcommittees and the Aboriginal Traditional Knowledge subcommittee. The Committee meets to consider status reports on candidate species.
Definitions (2016)
- Wildlife species
- A species, subspecies, variety, or geographically or genetically distinct population of animal, plant or other organism, other than a bacterium or virus, that is wild by nature and is either native to Canada or has extended its range into Canada without human intervention and has been present in Canada for at least 50 years.
- Extinct (X)
- A wildlife species that no longer exists.
- Extirpated (XT)
- A wildlife species no longer existing in the wild in Canada, but occurring elsewhere.
- Endangered (E)
- A wildlife species facing imminent extirpation or extinction.
- Threatened (T)
- A wildlife species likely to become endangered if limiting factors are not reversed.
-
Special concern (SC)
(Note: Formerly described as “Vulnerable” from 1990 to 1999, or “Rare” prior to 1990.) - A wildlife species that may become a threatened or an endangered species because of a combination of biological characteristics and identified threats.
-
Not at risk (NAR)
(Note: Formerly described as “Not In Any Category”, or “No Designation Required.”) - A wildlife species that has been evaluated and found to be not at risk of extinction given the current circumstances.
-
Data deficient (DD)
(Note: Formerly described as “Indeterminate” from 1994 to 1999 or “ISIBD” [insufficient scientific information on which to base a designation] prior to 1994. Definition of the [DD] category revised in 2006.) - A category that applies when the available information is insufficient (a) to resolve a species’ eligibility for assessment or (b) to permit an assessment of the species’ risk of extinction.
The Canadian Wildlife Service, Environment and Climate Change Canada, provides full administrative and financial support to the COSEWIC Secretariat.
Wildlife species description and significance
Name and classification
Phylum: Arthropoda – the arthropods
Subphylum: Hexapoda – the hexapods
Class: Insecta – the insects
Order: Orthoptera – the grasshoppers, crickets, katydids
Suborder: Caelifera – the grasshoppers
Superfamily: Acridoidea
Family: Acrididae – short-horned grasshoppers
Subfamily: Melanoplinae – spur-throated grasshoppers [in part; often considered part of the subfamily Cyrtacanthacridinae]
Genus: Melanoplus
Species: Melanoplus madeleineaeVickery and Kevan 1978
English Name: Magdalen Islands Grasshopper
French Name: Criquet des Îles-de-la-Madeleine
Taxonomic Background:
Melanoplus madeleineae, henceforth referred to as the Magdalen Islands Grasshopper, was described from specimens collected on the Magdalen Islands, Québec in 1975, and historical specimens dating back as far as 1912 (Vickery and Kevan 1978). The species was described by Vickery and Kevan (1978) based on diagnostic morphological characters. Recent molecular analysis supports the taxonomic validity of this species (Chapco and Litzenberger 2002). The Magdalen Islands Grasshopper is closely related (i.e., sister taxon) to the Northern Spur-throat Grasshopper (M. borealis), although the latter species does not occur on the Magdalen Islands (Kevan and Vickery 1978; Vickery and Kevan 1985; Chapco and Litzenberger 2002).
The Magdalen Islands Grasshopper belongs to the short-horned grasshopper family (Family Acrididae), which is characterized by their short antennae. This family contains three subfamilies: Melanoplinae, Gomphocerinae and Oedipodinae. The Magdalen Islands Grasshopper is part of the large subfamily Melanoplinae (spur-throated grasshoppers), which are primarily forb feeding specialists, and so named for the spur (or prosternal spine) located ventrally between the front legs. The subfamily Melanoplinae includes the most commonly encountered grasshoppers, with the genus Melanopluscontaining 239 species in North America (Arnett 2000), and 42 species within Canada (Vickery and Kevan 1985). The other two subfamilies are readily distinguishable morphologically and biologically from Melanoplinae.
Morphological description
The Magdalen Islands Grasshopper develops and grows by gradual metamorphosis during which there are three life stages: egg, nymph (immature forms) and adult. Gradual metamorphosis means the nymphs look similar to adults, progressively getting larger with each successive moult.
Adults:
The Magdalen Islands Grasshopper (Figures 1 and 2) is considered large for grasshoppers (female: 24 – 29 mm body length; male: 21 – 25.5 mm body length). Like other spur-throated grasshoppers, the species has an obvious spur located ventrally between the front legs. The antennae are typically threadlike (i.e., not flattened or sword-shaped). The head of most spur-throated grasshoppers, including the Magdalen Islands Grasshopper, is vertically oriented (i.e., not strongly slanted posteriorly from top to bottom like in the slant-faced grasshoppers) and not especially large in proportion to the body. The forewings are slightly pigmented, but lack the transverse bands common in the band-winged grasshoppers. The hindwings are not pigmented. The flying ability of spur-throated grasshoppers varies greatly, even within a single genus and the Magdalen Islands Grasshopper is likely not a strong flier (Vickery and Kevan 1985). The species is considered soft-bodied when compared with the sclerotized bodies of other grasshoppers.
Distinguishing features of the Magdalen Islands Grasshopper include the crimson colour of the lower edge of the hind femur, the non-banded outer face of the hind femur, and the golden-brown markings near the eyes (Vickery and Kevan 1978, 1985) (Figures 1 and 2). The colour and morphology of the Magdalen Islands Grasshopper closely resembles the closely related taxon, the Northern Spur-throat Grasshopper, although the former is slightly larger and the males have differently shaped genitalia (Vickery and Kevan 1985). Northern Spur-throat Grasshopper is also not present on Magdalen Islands (Vickery and Kevan 1985).

Long description for Figure 1
Photo of a female Magdalen Islands Grasshopper specimen. Distinguishing features of the Magdalen Islands Grasshopper include the crimson colour of the lower edge of the hind femur, the non-banded outer face of the hind femur, and the golden-brown markings near the eyes.

Long description for Figure 2
Photo of a male Magdalen Islands Grasshopper specimen. Distinguishing features of the Magdalen Islands Grasshopper include the crimson colour of the lower edge of the hind femur, the non-banded outer face of the hind femur, and the golden-brown markings near the eyes.
Immature stages:
Nymph stages (immature form) look similar to adults but lack fully developed wings, genitalia and body scleritization, which enable one to confirm taxonomic verification. The immature stages of Magdalen Islands Grasshopper have not been described nor are the number of immature moults from egg to adult known. However, for the related Northern Spur-throat Grasshopper, nymphs mature through five instars, so the same is presumed for Magdalen Islands Grasshopper. Based on the description of the Northern Spur-throat Grasshopper, the first instar is mainly dark except for a narrow, mid-dorsal brown band and a light crescent on the side of the head and pronotum. The general body colour of the second to fifth instars is typically greenish or brown. Nymphs were collected during the 2015 fieldwork in preparation for this status report.
Eggs:
The eggs of Magdalen Islands Grasshopper have not been described. However, for the related Northern Spur-throat Grasshopper, eggs are elongate and usually brown in colour, laid in pods of 30 – 50.
Population spatial structure and variability
The known range of the Magdalen Islands Grasshopper is restricted to the Magdalen Islands, a small geographic area that includes numerous islands and islets. The main chain of the islands (Île du Havre Aubert, Île du Cap aux Meules, Île du Havre aux Maisons, Grosse Île, Île de l’Est and Île de la Grande Entrée) are connected by sand dunes or other small sections of land.
Studies have supported the endemism of the Magdalen Islands Grasshopper based on its genetic distinctiveness from its sister taxon, the Northern Spur-throat Grasshopper (Chapco and Litzenberger 2002), which is not known from the islands (Vickery and Kevan 1985). The geological event leading to the separation of these species is estimated to have occurred approximately 85,000 to 11,000 years ago (Chapco and Litzenberger 2002).
Little is known of the spatial structure of Magdalen Islands Grasshopper subpopulations. The species is not considered a strong flier (Vickery and Kevan 1985) and its ability to migrate across islands is likely limited. However, this species was recently recorded from Île d’Entrée, which is 4.8 km from the main island group (Catling et al. 2013). It is likely this species has always been present on this island, but not detected.
Designatable units
The Magdalen Islands Grasshopper is being assessed as one designatable unit. The species is endemic to and has been recorded from seven of the fourteen islands that make up the Magdalen Islands. There are no data available on discreteness or evolutionary significance among the subpopulations on these seven islands. The species occurs within the COSEWIC Atlantic Ecological Area.
Special significance
The Magdalen Islands Grasshopper has been rarely collected (Catling et al. 2013) and is endemic to the Magdalen Islands (Vickery and Kevan 1978, 1985; Catling et al. 2013). This species is believed to represent a unique taxon derived from the Northern Spur-throat Grasshopper during isolation during the Wisconsin glaciation period (Cameron 1962; see Vickery and Kevan 1985).
Though part of the Scotian continental shelf in northeastern North America, the Magdalen Islands have been isolated from the mainland at least since the retreat of the last Wisconsin ice sheet approximately 85,000 to 11,000 years ago (Cameron 1962). Presumably, with lower sea levels in the past prior to the retreat, the Northern Spur-throat Grasshopper’s range would have extended onto vegetated landmasses connected to the continent. Rising sea level associated with the retreat of the ice sheet would have isolated populations of the Northern Spur-throat Grasshopper remaining on higher elevation landmasses separated from lower areas on the Scotian Shelf, ultimately promoting speciation with no gene flow with populations on the continent. Molecular evidence supports a very close affinity of the Magdalen Islands Grasshopper to mainland populations of the Northern Spur-throat Grasshopper. Hamilton (2002), Fortin and Larocque (2003) and Rémillard et al. (2016) provide detailed accounts of the glaciation history of the Magdalen Islands. Hamilton (2002) also discusses isolation and speciation of the insect fauna.
Distribution
Global and Canadian range
The global and Canadian distribution of the Magdalen Islands Grasshopper is restricted to the Magdalen Islands within the Gulf of Saint Lawrence, Québec, Canada (Figure 3). This archipelago is composed of eight main islands and several other small uninhabited islands with a land area of 205 km2 (totalling 14 islands and islets). Six of the main islands (Île du Havre Aubert, Île du Cap aux Meules, Grosse Île, Île du Havre aux Maisons, Pointe aux Loups, and Île de la Grande Entrée) are connected by sand bars, and the Magdalen Islands Grasshopper likely occurs on all of these islands. Two large islands are not connected, and are separated from the main island chain by ocean. Île d’Entrée is 4.8 km from the southeastern tip of the main group, and is known to support a subpopulation of the Magdalen Islands Grasshopper (Catling et al. 2013); Brion Island is approximately 13 km from Grosse Île in the north, but has not been surveyed for Magdalen Islands Grasshopper, nor have several other tiny uninhabited islands that are also considered part of the archipelago: Rocher aux Oiseaux, Île aux Loups Marins, Île Paquet and Rocher du Corps-Morts.

Long description for Figure 3
Map of North America showing the location of the Magdalen Islands within the Gulf of Saint Lawrence, Quebec.

Long description for Figure 4
Map showing the location of the two islands (Île du Havre aux Maisons and Île d’Entrée) in the Magdalen Islands group where the Magdalen Islands Grasshopper was confirmed most recently.

Long description for Figure 5
Map illustrating the maximum extent of occurrence of the Magdalen Islands Grasshopper and grid cells used to calculate index of area of occupancy.
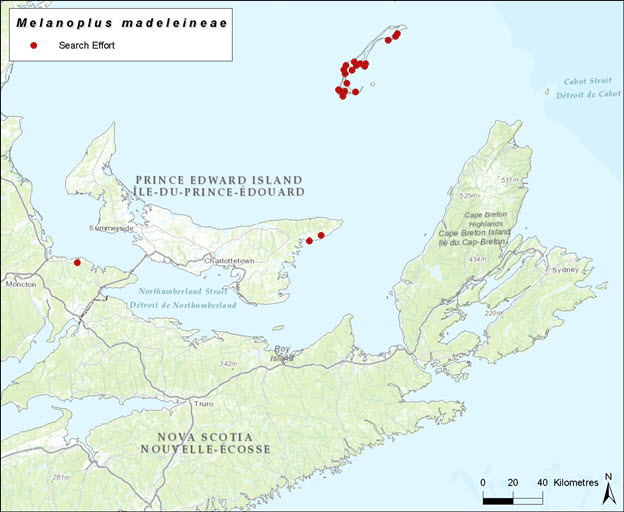
Long description for Figure 6
Map showing localities (in Quebec [Magdalen Islands], Prince Edward Island, and New Brunswick) searched for the Magdalen Islands Grasshopper in 2015 in preparation for this report.

Long description for Figure 7
Photo showing Magdalen Islands Grasshopper habitat consisting of open grassy maritime meadow. Flowering plants include Oxeye Daisy (Eucanthemum vulgare).

Long description for Figure 8
Photo showing Magdalen Islands Grasshopper habitat consisting of open grassy maritime meadow.

Long description for Figure 9
Photo of open grassy maritime meadow where immature Magdalen Islands Grasshoppers were collected and barcoded.

Long description for Figure 10
Photo of open grassy maritime meadow where immature Magdalen Islands Grasshoppers were collected and barcoded.

Long description for Figure 11
Photo of open grassy maritime meadow where immature Magdalen Islands Grasshoppers were collected and barcoded.

Long description for Figure 12
Map of the Magdalen Islands showing two sectors of habitat that could be flooded because of rising sea levels due to climate change.
Extent of occurrence and area of occupancy
The land area of the Magdalen Islands is 205.5 km2. The maximum extent of occurrence (EOO) for the Magdalen Islands Grasshopper is approximately 1021 km2, based on a minimum convex polygon that encompasses all of the Magdalen Islands. The index of area of occupancy (IAO) using a grid of 2 km X 2 km squares is 568 km2. The species does not occupy all habitats included in this spatial calculation, such as dense woodlands or beachside sandy areas, and the open unsuitable ocean habitat, and therefore the biological area of occupancy is likely much smaller.
Search effort
The Magdalen Islands Grasshopper is considered endemic to the Magdalen Islands based on the examination of many Melanoplus specimens from across Canada (see Vickery and Kevin 1978; Vickery and Kevin 1985). There are approximately 80 museum specimens of the Magdalen Islands Grasshopper, with collection dates ranging from 1912 to 2015 (Table 1) (including specimens reported from Vickery and Kevan (1978), Chapco and Litzenberger (2002), and Catling et al. (2013).
| Collection Site Name | No. of Specimens | Sex | Status | Collection Date | Collector(s) | Museum* | Reference |
|---|---|---|---|---|---|---|---|
| Île du Havre aux Maisons | 1 | - | - | August 11, 2010 | P.M. Catling, B. Kostiuk | CNC | Catling et al. 2013 |
| Île du Havre aux Maisons | 1 | - | - | August 13, 2010 | P.M. Catling, B. Kostiuk | CNC | Catling et al. 2013 |
| Île d’Entrée | 1 | - | - | August 14, 2010 | P.M. Catling, B. Kostiuk | CNC | Catling et al. 2013 |
| Île du Havre aux Maisons, Havre aux Maisons Parish, between Havre aux Maisons and Pointe Basse | 1 | ♂ | Holotype | September 9, 1975 | K.E. Kevan | LM | Vickery and Kevan 1978 |
| Île de la Grande Entrée, rise overlooking Bassin de l’Île Boudreau | 1 | ♀ | Allotype | September 9, 1975 | D.K. McE. Kevan | LM | Vickery and Kevan 1978 |
| Île du Havre aux Maisons, Havre aux Maisons Parish, between Havre aux Maison and Pointe Basse | 2 | ♂ | Paratype | September 8, 1975 | D.J. Luckin | LM | Vickery and Kevan 1978 |
| Île du Havre aux Maisons, Havre aux Maisons Parish, between Havre aux Maisons and Pointe Basse | 5 | ♀ | Paratype | September 8, 1975 | D.J. Luckin | LM | Vickery and Kevan 1978 |
| Île du Havre aux Maisons, Havre aux Maisons Parish, between Havre aux Maisons and Pointe Basse | 1 | ♂ | Paratype | September 8, 1975 | K.E. Kevan | LM | Vickery and Kevan 1978 |
| Île du Havre aux Maisons, Havre aux Maisons Parish, between Havre aux Maisons and Pointe Basse | 1 | ♂ | Paratype | September 8, 1975 | D.K. Kevan | LM | Vickery and Kevan 1978 |
| Île du Havre aux Maisons, Havre aux Maisons Parish, between Havre aux Maisons and Pointe Basse | 1 | ♀ | Paratype | September 8, 1975 | D.K. Kevan | LM | Vickery and Kevan 1978 |
| Southeast of town of Havre aux Maisons | 1 | ♀ | Paratype | September 11, 1975 | D.K. Kevan | LM | Vickery and Kevan 1978 |
| Cap Alright | 1 | ♀ | Paratype | September 9, 1975 | D.J. Luckin | LM | Vickery and Kevan 1978 |
| Île du Cap aux Meules, Fatima Parish, Chemin Patton | 1 | ♂ | Paratype | September 11, 1975 | D.J. Luckin | LM | Vickery and Kevan 1978 |
| Île du Cap aux Meules, Fatima Parish, Chemin Patton | 1 | ♀ | Paratype | September 11, 1975 | D.K. Kevan | LM | Vickery and Kevan 1978 |
| Île du Cap aux Meules, Fatima Parish, Chemin Patton | 1 | ♀ | Paratype | September 11, 1975 | K.E. Kevan | LM | Vickery and Kevan 1978 |
| Île du Havre Aubert, Parish | 2 | ♀ | Paratype | September 10, 1975 | D.K. Kevan | LM | Vickery and Kevan 1978 |
| Île du Havre Aubert, Cap Gridley | 1 | ♀ | Paratype | September 10, 1975 | D.K. Kevan | LM | Vickery and Kevan 1978 |
| South of Vigneau | 1 | ♀ | Paratype | September 10, 1975 | D.K. Kevan | LM | Vickery and Kevan 1978 |
| Île aux Loups, southwest of Pointe aux Loups | 1 | ♀ | Paratype | September 9, 1975 | D.K. Kevan | LM | Vickery and Kevan 1978 |
| “Îles de la Madeleine” | 1 | ♀ | Paratype | July 29, 1952 | J.R. Beaudry | Université de Montréal | Vickery and Kevan 1978 |
| “Îles de la Madeleine”, Août | 1 | ♀ | Paratype | "1952" | J.R. Beaudry | Université de Montréal | Vickery and Kevan 1978 |
| La Vernière | 1 | ♂ | Paratype | July 26, 1952 | J.R. Beaudry | Université de Montréal | Vickery and Kevan 1978 |
| La Vernière | 1 | ♀ | Paratype | July 26, 1952 | J.R. Beaudry | Université de Montréal | Vickery and Kevan 1978 |
| La Vernière | 1 | ♀ | Paratype | August 5, 1952 | J.R. Beaudry | Université de Montréal | Vickery and Kevan 1978 |
| Île du Cap aux Meules | 2 | ♂ | Paratype | July 29, 1952 | J.R. Beaudry | Université de Montréal | Vickery and Kevan 1978 |
| Étang du Nord | 1 | ♂ | Paratype | July 29, 1952 | J.R. Beaudry | Université de Montréal | Vickery and Kevan 1978 |
| Étang du Nord | 2 | ♀ | Paratype | July 29, 1952 | J.R. Beaudry | Université de Montréal | Vickery and Kevan 1978 |
| Île aux Loups [Wolfe I.] | 1 | ♀ | Paratype | August 21, 1912 | Bayard Long | Academy of Natural Sciences of Philadelphia | Vickery and Kevan 1978 |
| Île de la Grande Entrée [Coffin I.] | 1 | ♀ | Paratype | August 17, 1912 | Bayard Long | Academy of Natural Sciences of Philadelphia | Vickery and Kevan 1978 |
| Île du Havre aux Maisons [Alright I.] | 1 | ♂ | Paratype | August 21, 1912 | Bayard Long | Academy of Natural Sciences of Philadelphia | Vickery and Kevan 1978 |
| Île du Havre Aubert [Basin I.] | 1 | ♂ | Paratype | “1912” | Bayard Long | Academy of Natural Sciences of Philadelphia | Vickery and Kevan 1978 |
| Île du Cap aux Meules [Grindstone I.] | 1 | ♂ | Paratype | August 23, 1912 | Bayard Long | Academy of Natural Sciences of Philadelphia | Vickery and Kevan 1978 |
| Île de la Grande Entrée [Coffin I.] | 1 | ♀ | Paratype | August 17, 1912 | Bayard Long | Royal Ontario Museum | Vickery and Kevan 1978 |
| Île du Cap aux Meules [Grindstone I.] | 1 | ♂ | Paratype | August 23, 1912 | Bayard Long | Royal Ontario Museum | Vickery and Kevan 1978 |
| Île du Havre aux Maisons; beachside meadow habitat | Need to finalize | immature | Genbank #? | July 7, 2015 | C. Sheffield and J. Heron | RSM | Sheffield and Heron pers. data |
| Chemin de la Belle Anse off Chemin des Caps | Need to finalize | immature | Genbank #? | July 10, 2015 | C. Sheffield and J. Heron | RSM | Sheffield and Heron pers. data |
CNC = Canadian National Collection of Insects, Arachnids and Nematodes, Ottawa, ON.
LM = Lyman Museum, McGill University, Montréal, QC.
RSM = Royal Saskatchewan Museum, Regina, SK
| Month | Day | Year | Search Effort (minutes) for two people | Province | Location | Habitat Description | Collected Grass-hoppers |
|---|---|---|---|---|---|---|---|
| July | 6 | 2015 | 70 | NB | Exit 53, Highway 15, Cap Pelé exit | Roadside flower verge, approximately 20 - 30 metres wide, moist with abundant flowering plants. | - |
| July | 6 | 2015 | 90 | PE | Souris, outside of; near Lighthouse and Beach Motel | Grassy and wetland area adjacent to beachside; sandy soil | - |
| July | 7 | 2015 | 18 | PE | Souris, outside of; roadside ditch area | Roadside flower verge with extensive lupine patches | - |
| July | 8 | 2015 | 170 | QC: Magdalen Islands | Grosse Île et Grande Entrée | tip of the spit; meadow and sand dune grass | no |
| July | 8 | 2015 | 90 | QC: Magdalen Islands | spruce-fir woodlands at roadside | spruce-fir woodlands at the roadside | - |
| July | 9 | 2015 | 270 | QC: Magdalen Islands | Étang du Nord; municipal property | wetland adjacent to sand dunes (ocean) and pond (freshwater); collected in two habitats - along sand dune interface and within a wetland/boggy habitat. | yes |
| July | 9 | 2015 | 220 | QC: Magdalen Islands | Réserve naturelle de la Grande-Demoiselle - Paysage Culturel Site 12 | - | yes (immature) |
| July | 9 | 2015 | 30 | QC: Magdalen Islands | Open grassy area adjacent to a parking lot in an urban centre | grassy verge adjacent to ravine with flowing creek; invasive plants and weeds | yes |
| July | 9 | 2015 | 90 | QC: Magdalen Islands | Beach, roadside pull-out | beach/grass roadside pull-out adjacent to a sandy dune along the seaside; not hilly (lowlands) | yes (immature) |
| July | 10 | 2015 | 70 | QC: Magdalen Islands | Pointe Basse Cap Alright | flat meadow habitat (< 5% slope; 26m elevation); low maritime meadow with shorter vegetation intermixed with longer grasses and thicker areas dominated by milk vetch. | - |
| July | 10 | 2015 | 20 | QC: Magdalen Islands | Roadside | grassy roadside with lady beetles | no |
| July | 10 | 2015 | 70 | QC: Magdalen Islands | grassy hilly meadow | grassy hilly meadow adjacent to roadside; 54m elevation; some exposed rock | yes (immature) |
| July | 10 | 2015 | 210 | QC: Magdalen Islands | Cap à Fidèle, site de la Côte - Site 4 | maritime meadow - good bee/hymenoptera site; highly disturbed habitats with weedy patches adjacent to a wharf with commercial development (boat moorage) and restaurant; frequent traffic and parking | yes |
| July | 10 | 2015 | 90 | QC: Magdalen Islands | Picnic area at Chemin de la Belle-Anse off Ch. Des Caps Road | juniper meadow; very short vegetation and grasses; scattered vetch, buttercup, red clover, sedge, strawberry | - |
| July | 10 | 2015 | 120 | QC: Magdalen Islands | Pointe à Frank- Site 6 | near ocean | - |
| July | 10 | 2015 | Light trapping | QC: Magdalen Islands | Pointe de l’Est National Wildlife Area CWS property | sandy open dune habitat with heather and other dune plants | - |
| July | 8 - 11 | 2015 | Ongoing over 7 days | QC: Magdalen Islands | Home of 47 Chemin vigneau, Havre-aux-Maisons | Grassy backyard with lupines, and other wildflowers | no |
| July | 11 | 2015 | 108 | QC: Magdalen Islands | Baie du Havre aux Basques | beachside along roadway; pullout parking lot | yes (not in sedge areas where the habitat appears to be flooded; in the open sandy higher grassy areas that are not likely flooded) |
| July | 11 | 2015 | 120 | QC: Magdalen Islands | Cap au Sud; Lighthouse private property | maritime meadow adjacent to ocean (steep cliff drop-off > 40m) with a lighthouse. | yes |
| July | 11 | 2015 | 60 | QC: Magdalen Islands | La dune de l’ouest; Veille sur les Îsles | sand dunes next to ocean and adjacent to a large wetland/marsh; cranberry bog; flat habitat | yes (few) |
| July | 11 | 2015 | 42 | QC: Magdalen Islands | Chemin de la Montagne/Buttes intersection; roadside gravel areas | forest habitat with Abies spp. | - |
| July | 11 | 2015 | 78 | QC: Magdalen Islands | Adjacent to radio tower; Intersection of Montagne and Chemin du Grand-Pré roads | open meadow habitat with some conifer trees and mixed forest | yes |
| July | 11 | 2015 | 96 | QC: Magdalen Islands | Cap à Fidèle site de la Cote Site 4; caught Methocha! | federal Canada wharf area; maritime meadow - good bee/hymenoptera site; highly disturbed habitats with weedy patches adjacent to a wharf with commercial development (boat moorage) and restaurant; frequent traffic and parking | yes |
| July | 12 | 2015 | 120 | QC: Magdalen Islands | Pointe de l'Est National Wildlife Area CWS property | sandy open dune habitat with heather and other dune plants | - |
Table 3. The International Union for Conservation of Nature – Conservation Measures Partnership (IUCN-CMP) threats calculator outputs for the Magdalen Islands Grasshopper.
- Species or ecosystem scientific name:
- Magdalen Islands Grasshopper (Melanoplus madeleineae)
- Date:
- 2015-12-08
- Assessor(s):
- Cory Sheffield and Jennifer Heron (report writers); Angèle Cyr (COSEWIC Secretariat); Paul Grant (Co-Chair Arthropods SSC); Nathalie Desrosiers (Ministère des Forêts, de la Faune et des Parcs); Mathieu Morin (Ministère des Forêts, de la Faune et des Parcs - Manager Magdalen Islands); James Miskelly (Arthropods SSC member), John Klymko (Arthropods SSC member), Jessica Linton (Arthropods SSC member)
| Threat impact | Threat impact (descriptions) | Level 1 Threat impact counts: high range |
Level 1 Threat impact counts: low range |
|---|---|---|---|
| A | Very high | 0 | 0 |
| B | High | 0 | 0 |
| C | Medium | 0 | 0 |
| D | Low | 3 | 3 |
| - | Calculated overall threat impact: | Low | Low |
- Assigned overall threat impact:
- D = Low
| # | Threat | Impact (calculated) |
Scope (next 10 Yrs) |
Severity (10 Yrs or 3 Gen.) |
Timing | Comments |
|---|---|---|---|---|---|---|
| 1 | Residential and commercial development | Negligible | Negligible (<1%) | Moderate (11-30%) | High (Continuing) | - |
| 1.1 | Housing and urban areas | Negligible | Negligible (<1%) | Moderate (11-30%) | High (Continuing) | There is minimal new housing development within the Magdalen Islands. Most lots are large and the foot print of a house within a parcel of property is only a small portion of the lot size. There are municipal bylaws in place to limit property development within the rural areas of the islands. |
| 2 | Agriculture and aquaculture | Unknown | Small (1-10%) | Unknown | High (Continuing) | - |
| 2.1 | Annual and perennial non-timber crops | Unknown | Small (1-10%) | Unknown | High (Continuing) | The use of semi-natural agricultural areas, such as hay fields, by the Magdalen Islands Grasshopper is unknown. Creation of open habitats may have had a positive effect on the grasshopper. There are 600 ha of hay in production on the Magdalen Islands harvested 1-2 times per year (Morin pers. comm. 2015). Impacts of hay crop harvest on the closely related Northern Spur-throat Grasshopper suggest this species is tolerant of haying. |
| 2.2 | Wood and pulp plantations | - | - | - | - | Not applicable. The grasshopper does not occur in wooded areas. |
| 2.3 | Livestock farming and ranching | Unknown | Small (1-10%) | Unknown | High (Continuing) | Applicable. There is some grazing in pastures on the island, but the severity of this impact is unknown. |
| 3 | Energy production and mining | Negligible | Negligible (<1%) | Extreme (71-100%) | Moderate (Possibly in the short term, < 10 yrs) | - |
| 3.3 | Renewable energy | Negligible | Negligible (<1%) | Extreme (71-100%) | Moderate (Possibly in the short term, < 10 yrs) | At present, there is no large large-scale solar and wind energy infrastructure or generation on the island. However, a proposal for the installation of three wind turbines is under consideration by the Québec government. The footprint of the planned wind turbine infrastructure is directly within the same habitat as an endangered plant, as well as the habitats of other species at risk. It is unknown if the grasshopper is within this same footprint. |
| 4 | Transportation and service corridors | Low | Small (1-10%) | Serious (31-70%) | High (Continuing) | - |
| 4.1 | Roads and railroads | Low | Small (1-10%) | Serious (31-70%) | High (Continuing) | Applicable. Potential grasshopper mortality from vehicles, particularly on dirt roads where grasshoppers may take rest and/or females oviposit. |
| 5 | Biological resource use | - | - | - | - | - |
| 6 | Human intrusions and disturbance | Low | Small (1-10%) | Moderate (11-30%) | High (Continuing) | - |
| 6.1 | Recreational activities | Low | Small (1-10%) | Moderate (11-30%) | High (Continuing) | Applicable. The Magdalen Islands are a popular tourist destination for recreational activities. The number of visitors has increased over the years (See Threats). |
| 7 | Natural system modifications | - | Not a Threat | Small (1-10%) | Neutral or Potential Benefit | - |
| 7.3 | Other ecosystem modifications | - | Not a Threat | Small (1-10%) | Neutral or Potential Benefit | Neutral or beneficial impact. There may be some dyke construction to prevent erosion. This would stabilize some of the areas and prevent habitat loss. |
| 8 | Invasive and other problematic species and genes | Low | Pervasive (71-100%) | Slight (1-10%) | High (Continuing) | - |
| 8.1 | Invasive non-native/alien species | Low | Pervasive (71-100%) | Slight (1-10%) | High (Continuing) | Applicable. There are many non-native predators that potentially consume grasshoppers. (See Threats) |
| 9 | Pollution | - | - | - | - | - |
| 10 | Geological events | Not calculated (unknown timing) | Small (1-10%) | Extreme (71-100%) | Unknown | - |
| 10.2 | Earthquakes/ tsunamis | Not calculated (unknown timing) | Small (1-10%) | Extreme (71-100%) | Unknown | Potentially applicable. The six major islands are connected by sand bars. Earthquakes and tsunamis could cause some short term damage to connecting habitats. |
| 11 | Climate change and severe weather | Unknown | Pervasive (71-100%) | Unknown | Moderate (Possibly in the short term, < 10 yrs) | - |
| 11.1 | Habitat shifting and alteration | Not Calculated (outside assessment timeframe) | Small (1-10%) | Unknown | Low (Possibly in the long term, >10 yrs) | Potential sea level rise may impact shoreline beach habitats. |
| 11.2 | Droughts | Not Calculated (outside assessment timeframe) | Pervasive (71-100%) | Unknown | Low (Possibly in the long term, >10 yrs) | Periodic extreme droughts may impact the forage vegetation available for the Magdalen Islands Grasshopper. In cases of extreme drought, it is possible dormant eggs may not hatch due to unsuitable weather conditions. This threat is unknown because the specific life history details of this species are unknown. |
| 11.3 | Temperature extremes | Unknown | Pervasive (71-100%) | Unknown | Moderate (Possibly in the short term, < 10 yrs) | Extreme cold temperatures could potentially impact the grasshopper directly (cause mortality) or indirectly by damaging plant forage abundance. Climate models for the Maritime regions suggest that the climate will become even cooler and wetter, which may put pressure on the physiological tolerances of grasshoppers on the islands. This threat is unknown. |
| 11.4 | Storms and flooding | Not Calculated (outside assessment timeframe) | Unknown | Unknown | Low (Possibly in the long term, >10 yrs) | The Magdalen Islands naturally experiences strong winds and large storms; which could cause erosion of habitat. |
Past entomological collection events on the Magdalen Islands have been infrequent, and the islands have received targeted surveys on only a few occasions and for a few other insect groups (e.g., Kevan and Vickery 1978; Hamilton 2002). Relatively few specimens of the Magdalen Islands Grasshopper have been collected (Table 1) (Catling et al. 2013). The species was described in 1978, and 32 specimens were included in the type series, deposited at the Lyman Entomological Museum (13 specimens), Canadian National Collection of Insects, Arachnids and Nematodes (CNC) (2 specimens), Université de Montréal (10 specimens), Academy of Natural Sciences of Philadelphia (5 specimens), and the Royal Ontario Museum (2 specimens) (Vickery and Kevan 1978). At the time of this report writing, there are approximately 80 specimens held at the Lyman Museum, some of these including material in the type series from other institutions.
Grasshopper surveys in 1999 and 2000 yielded eight additional specimens of the Magdalen Islands Grasshopper that were subsequently used in a genetic study of this and closely related species (see Chapco and Litzenberger 2002); however, at present it is unknown if this material was destroyed for that study. Catling et al. (2013) collected three additional specimens in 2010, two specimens shown in Figures 1 and 2.
Field surveys were undertaken during the preparation of this status report. Surveys from July 6 – 12, 2015 totalled 40 hours over 23 sites (20 in the Magdalen Islands). Surveys were completed by hand collection and sweep netting open grassy habitats, roadsides adjacent to agricultural areas, open sand and sparsely vegetated areas and open wooded areas. All grasshoppers collected were aspirated out of the sweep net and placed into 70% ethanol until processing.
Approximately 280 grasshoppers were collected across all survey sites in 2015. The number of grasshoppers collected at each survey site was counted. All specimens collected were immature, so it was not possible to obtain accurate identification using taxonomic keys which use adult characteristics. These immature specimens were separated into morphological species groups and subsamples of these were processed for DNA barcoding. DNA barcoding generates a short genetic sequence (i.e., DNA barcode) from a standard portion of a specimens’ mitochondrial genome and compares this “barcode” to that of other species or specimens in the Barcodes of Life Database (BOLD). The presumption is that each species has a unique DNA barcode sequence (traditionally, biological specimens are identified using morphological features). The standard barcode for almost all animal groups is a 648 base-pair region in the mitochondrial cytochrome c oxidase 1 gene (“CO1”). From the selected individual grasshopper specimens, a single leg (or portion of leg for larger specimens) was removed and sent to the Biodiversity Institute of Ontario in Guelph, Ontario for tissue processing and sequencing. DNA barcode sequences, specimen images and all collection data were uploaded to the Barcodes of Life Data (BOLD) system, project “Magdalen Islands Grasshopper” (MAGIG). In addition, two recently collected adult specimens (provided by Paul Catling) were also DNA barcoded; these specimens serve as a genetic standard DNA barcode for Magdalen Islands Grasshopper.
DNA barcoding does not replace traditional taxonomy, but can help verify identification and/or can serve a dual purpose to inform the need to invest time and resources into improving the taxonomy around a species group. It also is useful when associating immature specimens to the adults. For grasshoppers, the timing of surveys may lead to inaccurate information on the relative abundance, including for the Magdalen Islands Grasshopper. Previous surveys may have been past the peak of adult emergence (Catling et al. 2013). Thus, DNA barcoding can be used as a tool for surveys where it is difficult to estimate the peak of adult emergence.
Based on DNA barcoding results, at least seven of the 280 specimens collected in 2015 were of Magdalen Islands Grasshopper. Barcoded specimens of Magdalen Islands Grasshopper were found only on Cap aux Meules (cliffside; Chemin de la Belle Anse off Chemin des Caps) and Île du Havre aux Maisons (beachside) (Table 1). This represents less than 3% of the total grasshoppers collected, suggesting that the species is relatively rare. In comparison, the Migratory Grasshopper (Melanoplus sanguinipes) accounted for over 80% of all specimens barcoded. Intraspecific variation in CO1 is considered low at 0.16%. Several other species of Melanoplusgrasshoppers collected in 2015 were also barcoded from the Magdalen Islands, though due to low interspecies variation in CO1 between these and mainland species, and suspected taxonomic errors in the Barcodes of Life Data system reference library for Melanoplus specimens, accurate identification based on DNA barcodes alone could not be made for these specimens. However, it was possible to determine these specimens were not the Magdalen Islands Grasshopper.
Habitat
Habitat requirements
The low number of survey and collection events makes it difficult to accurately characterize the species’ habitat. Catling et al. (2013) described the collection sites, in general, as open meadow habitats with native flora including, but not limited to Carex, Juncus, Eupatorium species.
Foraging and cover habitat:
Collections to date have been in open densely vegetated meadows. In 2015, immature specimens were collected from two sites that broadened the habitat description to include predominant grassy areas with an abundance of non-native grasses and forbs. In 2015, immature specimens of Magdalen Islands Grasshopper were collected at two sites, both sites with different habitats (Table 1). The habitat at the first collection site, Cap aux Meules (Chemin de la Belle Anse off Chemin des Caps) is an expansive grassy meadow adjacent to a steep sandy cliffside (approximately 30 m above sea level). Île du Cap aux Meules habitat has well-drained sandy soils and a moderate diversity of low-lying forb type plants that included, Fall Dandelion (Leontodon autumnalis), sumac (Rhusspp.), Queen Anne’s Lace (Daucus carota), buttercup (Ranunculus spp.), strawberry (Fragaria spp.), Red Clover (Trifolium repria), White Clover (Trifolium repens), Oxeye Daisy (Leucanthemum vulgare), hawkweed (Hieracium spp.), Wild Caraway (Carum carvi), and numerous species of yellow composites including goatsbeard (Trapopogonspp.). Non-native Tufted Vetch (Vicia cracca) was also present.
The second 2015 collection site was on Île du Havre aux Maisons and at beachside less than 5 m above sea level, adjacent to dune habitats (above dunes and the high tide line) with forb vegetation. The site had some of the plants above, but was dominated by Beach Pea (Lathyrus japonicus), Maram Grass (Ammophila breviligulata) and other long grasses.
Oviposition sites and overwintering habitat:
The Magdalen Islands Grasshopper likely overwinters as an egg (Vickery and Kevin 1985) within the soft sandy substrate of the open densely vegetated and grassy areas. Magdalen Island soils have a large sand component, with gravel and sandstone sediments. Oviposition sites chosen by gravid females are known to contribute to offspring fitness in grasshoppers (Fielding 2011).
Unlikely habitat:
During 2015 surveys, the grasshopper was not recorded within foredune open sand and sparsely vegetated dune plant communities. The absence of the Magdalen Islands Grasshopper and other grasshoppers from this habitat can be indirectly measured through results from a study of Piping Plover (Charadrius melodus) diet on the Magdalen Islands (Shaffer and Laport 1994). The absence of this grasshopper, or any orthopteran, in the droppings of this bird suggests that a grasshopper diet is not common in the coastal feeding habitats of these shore birds (Shaffer and Laport 1994). However, the presence of fast flying insects such as tiger beetles (Subfamily Cicindelinae) in the diet of Piping Plovers (Shaffer and Laport 1994; Majka and Shaffer 2008) suggests that grasshoppers, especially slow-moving gravid females, would likely be predated upon. The Magdalen Islands Grasshopper has also not been recorded from dense woodlands.
Habitat trends
The flora and fauna of the Magdalen Islands are estimated to have been stable for the past 8,000 to 11,000 years (Hamilton 2002). Over geological time, the archipelago may once have been part of larger glacial refugia during the Wisconsinan ice age (Hamilton 2002). Much of the Magdalen Islands is now inhabited, though the resident human population has remained relatively stable at 12,000 – 14,000 since the 1960s (Fortin and Larocque 2003), though the population increases with the tourist season to 50,000 people during the summer months.
Historically, and based on archeological evidence, the Magdalen Islands were visited by Mi’kmaq peoples, probably to harvest the abundant Walrus (Odobenus rosmarus) subpopulation that once resided on the islands. Prior to permanent settlement, the vegetation of the Magdalen Islands was mostly dense coniferous forests (i.e., Balsam Fir, Abies balsamea, and spruce, Picea spp.), though these were likely dwarfed and twisted by the wind into “elfinwoods”. Interspersed among these forests were boggy areas and low sand ridges (Hamilton 2002). The six main islands of the archipelago have experienced deforestation starting in the late 1800s and early 1900s. At present, GoogleEarth imagery (2015) shows that much of the vegetation on Île du Havre Aubert, Île du Havre aux Maisons, Île du Cap aux Meules and Île d’Entrée have been cleared and are now open meadow or pasture habitats, with little conifer forest remaining. The Magdalen Islands Grasshopper has not been recorded from conifer forests, and therefore the increase in open, meadow habitats may have benefited the species. Subsequently, a portion of these cleared areas have been converted to large rural house lots (Hamilton 2002).
Biology
Information on the biology and natural history of the Magdalen Islands Grasshopper is summarized from general references on grasshopper biology (Preston-Mafham 1990), and the closely related Northern Spur-throat Grasshopper (Vickery and Kevan 1985).
Life cycle and reproduction
Magdalen Islands Grasshopper has a life cycle typical of the genus Melanoplus (Pladt 2002) and most other grasshoppers. The species overwinters as an egg (Vickery and Kevan 1985). The similar species, Northern Spur-throat Grasshopper, requires two years to break egg diapause (Pladt 2002; Fielding 2008), and this may also apply to the Magdalen Islands Grasshopper.
Like all insects with gradual metamorphosis, several nymphal stages occur, each moulting and becoming progressively larger and with more pronounced adult characteristics. The Northern Spur-throat Grasshopper has five nymphal moults, and this is likely for the Magdalen Islands Grasshopper. Newly hatched grasshopper larvae leave the soil and climb up nearby surfaces and shed their first skin, entering the first-instar nymphal stage. The first instar is very tiny (approximately 2 to 3 mm in length), and without apparent wingbuds. As the individuals progresses through successive nymphal (immature) stages, wingbuds start to develop. The size (and shape) of the wingbuds is typically indicative of the developmental stage.
The first-instar stage signifies the commencement of the feeding and growth stages, and they feed, grow and eventually moult within a few days. The second nymphal life stage has a slightly longer body, but wingbud development is minor, each consisting of two small flaps. Again, after a few days (duration depending on temperatures), moulting occurs with the resulting third instar being longer, and with more pronounced wingbud development (i.e., some venation is visible). The fourth instar nymph is larger still, with small triangular wingbuds that have clear ridges (veins) and that extend over the first segment of the abdomen. These nymphs have a heavier body. This stage is the main period of feeding for grasshoppers. The last immature stage (the fifth) occurs when the individual is nearly mature. The wingbuds are much longer and resemble the fully developed wing. This stage typically lasts about a week, after which the grasshopper will moult, with fully developed wings (these take a full day to harden). These individuals breed within a week. Once egg diapause ends, development to adult stage generally takes approximately one month depending on species, and local weather conditions.
Adult Magdalen Islands Grasshoppers are active from late July through mid-September (Vickery and Kevan 1985). In the Northern Spur-throat Grasshopper, females begin oviposition 8 to 10 days after mating (Pladt 2002) although this is unknown for the Magdalen Islands Grasshopper. Females lay eggs in pods of 30 – 50 eggs per pod at a depth of 1 – 3 cm within the substrate, or against stones, plant roots or similar microsites. Depending on species and geography, eggs hatch in the late spring, if the soil is warm, or into early summer (mid-June). Hatching within a subpopulation is not simultaneous, and may be spread out over a period of two or more weeks. With so few data, it is difficult to estimate the peak adult activity period for the species.
Physiology and adaptability
No data on the specific physiology and adaptability of Magdalen Islands Grasshopper has been reported. It is likely that subpopulations of Magdalen Islands Grasshopper would survive if established on coastal areas of the mainland, though the genetic closeness of this species with the Northern Spur-throat Grasshopper, from which it is geographically isolated, may mean that it could possibly hybridize, although Vickery and Kevan (1985) suggest this is unlikely due to differing male genitalia.
Dispersal and migration
Little is known about the dispersal and migratory ability of the Magdalen Islands Grasshopper, but the adults are not considered strong fliers (Vickery and Kevan 1985). It currently is known from the six main connected Magdalen Islands, and more recently from Île d’Entrée which is located 4 km from the main islands. The next nearest landmass is Prince Edward Island (105 km south) and dispersal to this landmass is considered unlikely. It is also unknown whether the adults or immature stages can migrate from one island to the other (even when connected by sandbars).
Interspecific interactions
The Magdalen Islands Grasshopper is subject to predation by a wide range of vertebrates and invertebrates (Preston-Mafham 1990), the importance of each taxa depending on the habitat and geographic location. Vertebrate predators are considered the major driving force for some of the chemical defences that have evolved within the Orthoptera (Preston-Mafham 1990). Thus one can assume that vertebrates are important predators in most terrestrial habitats.
The terrestrial vertebrate diversity on the Magdalen Islands is low and the impacts from vertebrate predation to the Magdalen Islands Grasshopper are unknown. The extent to which native mammals prey on the Magdalen Islands Grasshopper is unknown; only a few species of terrestrial mammals have been reported (Cameron 1962), two of which are native: Meadow Vole (Microtus pennsylvanicus), and Deer Mouse, (Peromyscus maniculatus). There is conflicting information on whether bats are considered native or non-native, though bat species in eastern Canada are not considered major predators of grasshoppers (Brigham pers. comm. 2015). The Meadow Vole is also probably not a major predator of grasshoppers. However, the Deer Mouse has a diet that includes grasshoppers. Red Foxes (Vulpes vulpes) are not considered native to the Magdalen Islands although confirmed records of their presence on the islands date back to the late 1800s (Cory 1878). Red Foxes are known predators of grasshoppers (Catling 1988; Read and Bowen 2001). There are no native amphibians or reptiles on the Magdalen islands.
Birds are the major predators of grasshoppers and approximately 320 bird species inhabit the Magdalen Islands; with such a large diversity there are many potential predators (Preston-Mafham 1990). No orthopterans were found in a study of the diet (i.e., based on dropping contents) of Piping Plovers on the Magdalen Islands (Shaffer and Laport 1994). These data also suggests grasshoppers may not be abundant in plover habitats. The diet of Piping Plovers certainly includes several highly mobile arthropods (Shaffer and Laport 1994; Majka and Shaffer 2008) suggesting that these birds would be capable of capturing immature and adult grasshoppers if they were present and abundant.
Magdalen Islands Grasshoppers have been recorded from regurgitated pellets of Whimbrel (Numenius phaeopus) on Grosse Île. Whimbrels and other curlews are generalist insect predators (Skeel and Mallory 1996) and are not specific to grasshoppers. However, grasshoppers may form a large part of an ephemeral food source for Whimbrels in grassy habitats such as the Canadian prairies (Ray Poulin, pers. comm. 2016).
There are numerous generalist arthropod predators of orthopterans, including spiders (Aranae), predatory beetles (e.g., tiger beetles and ground beetles (Coleoptera: Carabidae)), robber flies (Diptera: Asilidae), ants and many predatory wasps (Hymenoptera) (Preston-Mafham 1990). The larvae of the genus Epicauta in the blister beetle family (Meloidae) are specialized predators of grasshopper eggs (Pinto 1991). In some areas they are important, capable of destroying close to 10% of grasshopper egg pods (Parker and Wakeland 1957). Their importance as a predator of the Magdalen Islands Grasshopper is unknown as no Epicauta have been reported on the Magdalen Islands.
Specific interspecific associations with grasshoppers come from their parasites and parasitoids. Several groups of Diptera (families Sarcophagidae, Anthomyiidae, Tachinidae, Nemestrinidae) and Hymenoptera (e.g., Scelionidae) are parasitoids of grasshoppers (Smith 1940; Smith and Finlayson 1950; York and Prescott 1952; Rees 1985; Przybyszewski and Capinera 1991; Laws and Joern 2012).
Mites are also important parasites of grasshoppers (Branson 2003), and are considered potentially important for future pest management initiatives for problematic species (Belovsky et al. 1997). Mites are commonly found on adult grasshopper wings although appear to have little impact. Conversely, some mite species are predaceous on grasshopper eggs and may have an impact on grasshopper subpopulations (Belovsky et al. 1997). The impact of mites on most grasshopper species, including Magdalen Islands Grasshopper is not well documented.
Biron et al. (2005) report a juvenile hairworm (Nematomorpha) as a parasite of orthopterans which alters adult grasshopper behaviour to induce them to jump in to fresh water, thus killing them. A range of nematodes are known parasites of grasshoppers, some used as biological control agents (Baker and Capinera 1997). However, no specific documentation of these parasites is confirmed for Magdalen Islands Grasshopper.
Fungal and bacterial pathogens are important factors affecting Orthoptera (Bucher and Stephens 1957; Streett and McGuire 1990; Glogoza and Weiss 2010). Probably the most important of these pathogens is the fungal species Entomopthora grylli, which is apparently specialized for grasshoppers and crickets (Pickford and Riegert 1964; Erlandson et al. 1988). The nymphal stages appear to be more susceptible to fungal infection than adults (Kistner and Belovsky 2013). Another fungal pathogen of potential importance is Nosema locustae (Ewen and Mukerji 1980). However, the status of these pathogens in Magdalen Islands Grasshopper subpopulations is undocumented.
Population sizes and trends
Sampling effort and methods
Surveys for the Magdalen Islands Grasshopper have focused on recording new subpopulations, natural history and habitat information, and collecting material for genetic studies. Surveys have been primarily by wandering transects through suitable habitat and sweep-netting habitat for grasshoppers. Catling et al. (2013) reported on the difficulty of timing collection events to correspond with peak adult emergence, suggesting that detection success could vary considerably. Collection events occurred between July 8-15, when most of the grasshopper species on the island were immature (we collected 1st-5th instar stages). Due to problems inherent in identifying immature grasshoppers, DNA barcoding was used as a means to identify species collected at different sites and estimate abundance.
Abundance
There are few data from which to estimate the abundance of Magdalen Islands Grasshopper at extant sites. Most observations and collections are one or two individuals. Results from DNA barcoding suggest that Magdalen Islands Grasshopper is relatively rare compared to other species on the islands, perhaps accounting for fewer than 3% of all specimens of Melanoplus encountered.
Fluctuations and trends
The natural subpopulation fluctuations in grasshoppers are a result of factors such as parasites, predators, food abundance and the previous years’ weather. For some species, fluctuations in population size may also result from varying levels of egg survival based on oviposition site selection (Fielding 2011). There is insufficient information to estimate subpopulation fluctuations or trends specifically for the Magdalen Islands Grasshopper. However, the species may experience extreme fluctuations based on information from its sister taxon, the Northern Spur-throat Grasshopper, as well as other related Melanoplus grasshopper species (Mulkern 1980).
Rescue effect
The Magdalen Islands Grasshopper is endemic to the Magdalen Islands; rescue from outside Canada is therefore not possible.
Threats and limiting factors
The International Union for Conservation of Nature-Conservation Measures Partnership (2006) (IUCN-CMP) threats calculator was used to classify and list threats to the Magdalen Islands Grasshopper (Salafsky et al. 2008; Master et al. 2009). The overall Threat Impact for the species is Low.
Low impact threats consist of roads and roadways where vehicles may cause mortality to grasshoppers that either cross roads and/or try to oviposit within the substrate of dirt roads; recreational activities including all-terrain vehicle operations in meadow habitats and invasive and other problematic species which are known to prey upon grasshoppers.
Threat 4. Transportation & Service Corridors (low impact)
4.1 Roads and railroads
Vehicular traffic along roadways may trample and impact grasshopper subpopulations using habitats adjacent to these areas (Catling et al. 2013). Additionally, pregnant females are slow-moving and tend to hop to move from place to place, pausing between jumps during which time they are vulnerable to being killed on the road. The substrates of numerous less-used roadways on the Magdalen Islands have a high component of sand mixed with gravel. Females are known to oviposit within these soft substrates, making both the eggs and females more vulnerable to mortality from road traffic. Vehicular traffic is highest during the summer months, when the islands population increases from tourism.
Roadside expansion and widening of roadways is unlikely as much of this infrastructure has been in place for many decades. Road repairs following large storm or flooding events are ongoing.
Threat 6. Human Intrusions & Disturbance (low impact).
6.1 Recreational activities
The Magdalen Islands are a popular tourist destination during the warm summer months of June through September and the islands’ economy is tied to recreational tourism. Recreational tourism has increased substantially in the past forty years. In the 1970s tourist numbers were 9,000 to 18,000 visitors per year; 1990s 30,000 visitors per year; 2002 had 54,000 visitors; and 2003 – 2005 had greater than 50,000 visitors per year; and tourists numbered 62,200 visitors in 2012 (Tourism Îles de la Madeleine 2016b). As of 2012, international cruises started stopping in the Magdalen Islands, thus increasing the number of tourists to the islands each summer.
Much of the recreational activity occurs within the coastline beach ecosystems, although in some places access to beaches is through the open meadow habitats where the grasshopper potentially occurs. There is a large network of open meadow parks, lookouts and protected hillsides within the Magdalen Islands that are popular destinations for hiking and dog walking. Some of these habitats are not protected, and vulnerable to more intensive activities such as mountain biking. Trampling and increased foot traffic along paths at some sites may trample grasshoppers (Catling et al. 2013). The Northern Spur-throat Grasshopper is known to spend much of its adult life basking on bare ground and if it is revealed the Magdalen Islands Grasshopper has similar preferences, the impacts to local populations will be more than currently estimated (e.g., adults may use open trails and roadways for basking and/or as oviposition sites).
The overall impact from recreational activities at present is considered low; however, they could become a larger threat should tourism grow, especially tourism related to international cruise ships.
Threat 8. Invasive & Other Problematic Species, Genes & Diseases (low impact)
8.1 Invasive Non-Native/Alien Species
European Starling (Sturnus vulgaris) could be a low impact threat to the Magdalen Islands Grasshopper (Catling et al. 2013) as it is found in increasingly high numbers on the islands, and is known to eat grasshoppers and other insect prey (Wood 1973). Other starling species are known to be important predators of grasshoppers, and have even been suggested as potential biological control agents (Ji et al. 2008). The gradual spread and invasion of non-native plants may change the floral abundance and composition available as grasshopper forage.
Domesticated and/or feral cats (Felis catus) can also be important predators of grasshoppers (Errington 1936; McMurry and Sperry 1941; Konecny 1987; Catling 1988; Read and Bowen 2001) and the frequency of predation on grasshoppers by cats has been reported to increase in the summer months when grasshoppers are larger and more abundant (Molsher et al. 1999).
Other problematic mammals that have been introduced and have established on the Magdalen Islands include the Norway Rat (Rattus norvegicus) (Cameron 1962), Red Squirrels (Sciurus vulgaris) and Snowshoe Hares (Lepus americanus) (Dinets 2015), and Minks (Neovison vison) (Tourism Îles de la Madeleine 2016a). Coyotes (Canis latrans) were first observed on the islands in 2001 (Naughton 2012).
Invasive insects, such as the European Fire Ant (Myrmica rubra), could potentially impact the Magdalen Islands Grasshopper, by consuming eggs as well as adults when females are slow and heavy with eggs. The ant is known to outcompete and reduce invertebrate faunal assemblages within its immediate nest area and foraging territory. At present, the European Fire Ant is not recorded on the Magdalen Islands. There may be some potential for the introduction of the related Northern Spur-throat Grasshopper via human transport (e.g., through transport on vegetative cuttings). The introduction of this grasshopper would likely result in interbreeding, hybridization and competition with the Magdalen Islands Grasshopper. Additional surveys of the insect fauna at known grasshopper habitats, to monitor any changes, are needed.
Limiting factors
The limiting factors applicable to the Magdalen Islands Grasshopper include:
Limited dispersal capability:
Females are not strong fliers and their slower movements may make the species vulnerable to trampling and predation.
Short growing season:
The Magdalen Islands has a short growing season, with July and August temperature record highs of 29°C and 31°C and record lows of 7.5°C and 6.6°C respectively. Harsh weather limits the time available to complete the grasshopper’s life cycle, including time to disperse to new habitats.
Fluctuating weather patterns:
Grasshoppers rely on the ambient temperature to warm their bodies and allow movement and the cool and foggy summer weather can be suboptimal for grasshopper movement. Year to year variation in weather patterns could affect subpopulation levels due to decreased foraging activity in extreme conditions. Strong winds can also lead to increased erosion levels at sites with exposed soil (e.g., affecting grasshopper eggs) and limit grasshopper movement, foraging and mating.
Number of locations
It is difficult to estimate a number of locations for the Magdalen Islands Grasshopper. The species is slow-moving, although likely mobile enough to repopulate suitable habitats over time. The predominant and most serious, plausible threats to the species are 4.1 road mortality, 6.1 recreational activities that adversely impact grasshopper habitat or cause direct mortality, and 8.1 predation on grasshoppers. These threats vary across the Magdalen Islands, therefore, the number of locations is likely a range of 7 (islands with confirmed records) to 14 (all islands and islets in the archipelago).
Protection, status and ranks
Legal protection and status
There are currently no federal or provincial laws that specifically protect the Magdalen Islands Grasshopper, mitigate threats to grasshoppers or protect the species’ habitat.
There may be habitat and occurrences of the Magdalen Islands Grasshopper within Pointe de L’Est National Wildlife Area although this wildlife area has not been thoroughly inventoried for the species. If the species was recorded from the wildlife area, it would be protected under the federal Wildlife Area Regulations (C.R.C., c. 1609).
In Québec, this species is not listed under the “Loi sur les espèces menacées ou vulnerable” (RLRQ, c E-12.01) (LEMV) (Act respecting threatened or vulnerable species) (CQLR, c E-12.01) and is not afforded protection under the “Loi sur la conservation et la mise en valeur de la faune” (RLRQ, c. C- 61.1) (LCMVF) (Act respecting the conservation and development of wildlife) (CQLR, c. C-61.1). Also, this species is not integrated on the Liste des espèces susceptibles d’être désignées menacées ou vulnérables (list of wildlife species likely to be designated threatened or vulnerable). This list is produced according to the “Loi sur les espèces menacées ou vulnérables” (RLRQ, c E-12.01) (LEMV) (Act respecting threatened or vulnerable species) (CQLR, c E-12.01).
Non-legal status and ranks
The conservation status ranks for the Magdalen Islands Grasshopper are:
- Global Status: G2 (Imperilled) (last reviewed December 2000)
- National Status: N2 (Imperilled) (last reviewed December 2000)
- Provincial Status: Québec S2 (Imperilled)
Habitat protection and ownership
The Magdalen Islands Conservation Society (SCÎM) is a non-government conservation organization (NGO) on the Magdalen Islands. This NGO works with private landowners on initiatives to protect species at risk and habitat and maintain the natural ecosystems of the Magdalen Islands (Déraspe pers. comm. 2015). To date, the organization has protected over 222 hectares of habitat across the archipelago (SCÎM 2015) although it is unknown if the grasshopper occurs in these habitats. Attention FragÎles is also a non-government conservation organization on the Magdalen Islands that works on collaborations with local citizens, planners and visitors to ensure environmental values on the islands are protected (Attention FragÎles 2015a).
Approximately 30% of the total land area of the Magdalen Islands is public land, including the major roadways and sand bars that link the islands together. It is unknown how much of this habitat is suitable or occupied by the Magdalen Islands Grasshopper.
Acknowledgements
The following people provided valuable information, advice, and knowledge: Paul Catling, Véronique Déraspe, Nathalie Desrosiers, Isabelle Gauthier, Mathieu Morin, John Klymko, Rémi Hebert and Andy Hamilton. The report writers wish to thank the Royal Saskatchewan Museum for providing time and resources to complete this report including partial funding for barcoding specimens.
The report writers wish to thank Marie Fortier for arranging permits to survey the Pointe de l’Est National Wildlife Area. Thanks are extended to curators and collections managers for enabling access to specimens. Paul Grant (Arthropods Specialist Subcommittee [SSC] Co-chair) is thanked for editorial and technical support, Angèle Cyr (COSEWIC Secretariat) for technical support, and Jenny Wu (COSEWIC Secretariat) for mapping support.
The cover photograph is by Paul Catling: Female Magdalen Islands Grasshopper photographed within open meadow habitat on Île du Havre aux Maisons in the Magdalen Islands, QC, with Île d’Entrée in the background on August 11, 2010. Photo originally published in Catling et al. (2013).
Authorities contacted
Catling, Paul. Research Scientist. Canadian National Collection of Insects, Arachnids and Nematodes, Agriculture and Agri-foods Canada. Ottawa, ON.
Déraspe, Véronique. Directrice, Société de conservation des Îles-de-la-Madeleine (SCÎM) 202-350, chemin Principal, Cap-aux-Meules, QC.
Desrosiers, Nathalie. Ministère des Forêts, de la Faune et des Parcs, QC.
Fortier, Marie. Protected Areas Specialist, Environment Canada, Québec, QC.
Gauthier, Isabelle. Coordonnatrice provinciale des espèces fauniques menacées et vulnérables, Ministère des Forêts, de la Faune et des Parcs, QC.
Hamilton, Andrew. Research Scientist. Canadian National Collection of Insects, Arachnids and Nematodes, Agriculture and Agri-foods Canada. Ottawa, ON.
Morin, Mathieu. Direction de la gestion de la faune de la Gaspésie-Îles-de-la-Madeleine, Ministère des Forêts, de la Faune et des Parcs, Sainte-Anne-des-Monts, QC.
Lonsdale, Owen. Collections Manager. Canadian National Collection of Insects, Arachnids and Nematodes, Agriculture and Agri-foods Canada. Ottawa, ON.
Wheeler, Terry. Lyman Entomological Museum, Department of Natural Resource Sciences McGill University, Macdonald Campus, Montréal, QC.
Information sources
Arnett, R.H. 2000. American Insects: A Handbook of the Insects of America North of Mexico. CRC Press.
Attention FragÎles. 2015a. Fighting together against erosion. When the citizens of the Magdalen Islands each do their part to slow erosion of the dunes [PDF, 4.04 Mb]. Available at [Accessed December 15, 2015].
Attention FragÎles. 2015b. Public consultations: Strategic environmental assessment on the whole of the hydrocarbons sector. Presentation to the Ministry of Energy and Mines. November 18, 2015. Consultations publiques : Évaluation environnementale stratégique sur l’ensemble de la filière des hydrocarbures. Présenté au ministère de l’Énergie et des Ressources naturelles [PDF, 890 Kb] [Accessed December 14, 2015] [Avaiable in French only]
Baker, G.L., and J.L. Capinera. 1997. Nematodes and nematomorphs as control agents of grasshoppers and locusts. Memoirs of the Entomological Society of Canada 171: 157-211.
Belovsky, G.E., D. Branson, J. Chase, J. Barker, and G. Hammond. 1997. Mites and Nematode Parasites of Grasshoppers. In: Grasshopper Integrated Pest Management User Handbook. USDA/APHIS/PPQ, Tech. Bull. 1809: 1-3, Boise, ID.
Bernatchez P., C. Fraser, S. Friesinger, Y. Jolivet, S. Dugas, S. Drejza, et A. Morissette A. 2008. Sensibilité des côtes et vulnérabilité des communautés du golfe du Saint-Laurent aux impacts des changements climatiques. Laboratoire de dynamique et de gestion intégrée des zones côtières, Université du Québec à Rimouski. Rapport de recherche remis au Consortium OURANOS et au FACC, 256 pages.
Biron, D.G., L. Marché, F. Ponton, H.D. Loxdale, N. Galéotti, L. Renault, C. Joly, and F. Thomas. 2005. Behavioural manipulation in a grasshopper harbouring hairworm: a proteomics approach. Proceedings of the Royal Society of London B: Biological Sciences 272(1577): 2117-2126.
Branson, D.H. 2003. Effects of a parasite mite on life-history variation in two grasshopper species. Evolutionary Ecology Research 5: 397-409.
Brigham, M. 2015. University of Regina. Personal communication to Cory Sheffield.
Bucher, G.E., and J.M. Stephens. 1957. A disease of grasshoppers caused by the bacterium Pseudomonas aeruginosa (Schroeter) Migula. Canadian Journal of Microbiology 3(4): 611-625.
Cameron, A.W. 1962. Mammalian zoogeography of the Magdalen Islands Archipelago, Québec. Journal of Mammalogy 43: 505-514.
Catling, P.C. 1988. Similarities and contrasts in the diets of foxes, Vulpes vulpes, and cats, Felis catus, relative to fluctuating prey populations and drought. Australian Wildlife Research 15: 307-317.
Catling, P.M., D.F. McAlpine, C.I. Adam, G. Belliveau, D. Doucet, A.D. Fairweather, and A.W. Thomas. 2013. New and noteworthy records of Orthoptera and allies in the Maritimes and the Îles-de-la-Madeleine, Québec. Canadian Field-Naturalist 127: 332-337.
Chapco, W., and G. Litzenberger. 2002. A molecular phylogenetic study of two relict species of melanopline grasshoppers. Genome 45: 313-318.
Dinets, V. 2015. Peterson Field Guide to Finding Mammals in North America. Houghton Mifflin Harcourt Publishing Company.
Erlandson, M. A., D.L. Johnson, and O.O. Olfert. 1988. Entomophaga grylli (Fresenius) infections in grasshopper (Orthoptera: Acrididae) populations in Saskatchewan and Alberta, 1985–1986. Canadian Entomologist 120: 205-209.
Errington, P.L. 1936. Notes on food habits of southern Wisconsin house cats. Journal of Mammalogy 17: 64-65.
Ewen, A.B., and M.K. Mukerji. 1980. Evaluation of Nosema locustae (Microsporidia) as a control agent of grasshopper populations in Saskatchewan. Journal of Invertebrate Pathology 35: 295-303.
Fielding, D.J., 2008. Diapause traits of Melanoplus sanguinipes and Melanoplus borealis (Orthoptera: Acrididae). Annals of the Entomological Society of America, 101: 439-448.
Fielding, D.J. 2011. Oviposition site selection by the grasshoppers Melanoplus borealisand M. sanguinipes (Orthoptera: Acrididae). Journal of Orthoptera Research, 20: 75-80.
Fielding, D.J. and L.S. Defoliart. 2010. Embryonic developmental rates of northern grasshoppers (Orthoptera: Acrididae): implications for climate change and habitat management. Environmental Entomology 39:1643-1651.
Fortin, J.C., and P. Larocque. 2003. Histoire des îles de la Madeleine. Laval University Press.
Glogoza, P.A., and M.J. Weiss. 2010. Grasshopper biology and management [PDF, 70.0 Kb]. Unpubished manuscript
Hamilton, K.G.A. 2002. Îles-de-la-Madeleine (Magdalen Is.): a glacial refugium for short-horned bugs (Homoptera: Auchenorrhyncha)? Le Naturaliste Canadien 126: 25-40.
Ji, R., S.J. Simpson, F. Yu, Q.X. He, and C.J. Yun. 2008. Diets of migratory rosy starlings (Passeriformes: Sturnidae) and their effects on grasshoppers: implications for a biological agent for insect pests. Biological Control 46: 547-551.
Kevan, D.K.M., and V.R. Vickery. 1978. The orthopteroid insects of the Magdalen Islands with notes from adjacent regions [Canada]. Annals of the Entomological Society of Québec 22: 193-204.
Kistner, E.J., and G.E. Belovsky. 2013. Susceptibility to disease across developmental stages: examining the effects of an entomopathogen on a grasshopper (Orthoptera: Acrididae) pest. Journal of Orthoptera Research 22: 73-77.
Konecny, M.J. 1987. Food habits and energetics of feral house cats in the Galapagos Islands. Oikos 50: 24-32.
Laws A.N., and A. Joern. 2012. Variable effects of dipteran parasitoids and management treatment on grasshopper fecundity in a tallgrass prairie. Bulletin of Entomological Research 102: 123-130.
LEMV (Liste des espèces susceptibles d’être désignées menacées ou vulnérables). Accessed September 8, 2016. [Avaiable in French only]
Magdalen Islands Conservation Society. 2015. [Accessed December 14, 2015].
Majka, C.G., and F. Shaffer. 2008. Beetles (Coleoptera) in the diet of Piping Plovers in the Iles de la Madeleine, Québec, Canada. Wader Study Group Bull. 115: 77–83.
McMurry, F.B., and C.C. Sperry. 1941. Food of feral house cats in Oklahoma, a progress report. Journal of Mammalogy 22: 185-190.
Molsher, R., A. Newsome, and C. Dickman. 1999. Feeding ecology and population dynamics of the feral cat (Felis catus) in relation to the availability of prey in central-eastern New South Wales. Wildlife Research 26: 593-607.
Mulkern, G.B. 1980. Population fluctuations and competitive relationships of grasshopper species (Orthoptera: Acrididae). Transactions of the American Entomological Society 106:1-41.
NatureServe. 2015. NatureServe Explorer: An online encyclopedia of life [web application]. Version 7.1. NatureServe, Arlington, Virginia. Available (Accessed: October 8, 2015).
Parker, J.R., and C.C. Wakeland. 1957. Grasshopper egg pods destroyed by larvae of bee flies, blister beetles, and ground beetles. US Department of Agriculture Technical Bulletin No. 1165.
Pickford, R., and P.W. Riegert. 1964. The fungus disease caused by Entomophthora grylli Fres., and its effects on grasshopper populations in Saskatchewan in 1963. Canadian Entomologist 96: 1158-1166.
Pinto, J.D. 1991. The taxonomy of North American Epicauta (Coleoptera: Meloidae), with a revision of the nominate subgenus and a survey of courtship behavior. University of California Publications in Entomology 110: 1-372.
Preston-Mafham, K. 1990. Grasshoppers and Mantids of the World. Facts on File, New York. 192 pp.
Przybyszewski, J., and J.L. Capinera. 1991. Patterns of parasitism among shortgrass prairie grasshopper (Orthoptera: Acrididae) populations. Journal of the Kansas Entomological Society 64: 5-17.
Read, J., and Z. Bowen. 2001. Population dynamics, diet and aspects of the biology of feral cats and foxes in arid South Australia. Wildlife Research 28: 195-203.
Rees, N.E. 1985. Suitability of selected North American grasshopper species as hosts for grasshopper parasites from Pakistan. Agriculture, Ecosystems & Environment 12: 157-163.
Rémillard, A. M., G. St-Onge, P. Bernatchez, J.P. Buylaert, A.S. Murray, and B. Vigneault, B. 2016. Chronology and stratigraphy of the Magdalen Islands archipelago from the last glaciation to the early Holocene: new insights into the glacial and sea-level history of eastern Canada. Boreas. 10.1111/bor.12179. ISSN 0300-9483.
Shaffer, F. and P. Laporte. 1994. Diet of Piping Plovers on the Magdalen Islands, Quebec. Wilson Bulletin 106: 531-536.
Skeel, M.A., and E.P. Mallory. 1996. Whimbrel (Numenius phaeopus). In The Birds of North America, No. 219 (A. Poole and F. Gill, eds.). The Academy of Natural Sciences, Philadelphia, PA, and the American Ornithologists' Union, Washington, D.C.
Smith, C.W. 1940. An Exchange of Grasshopper Parasites between Argentina and Canada with Notes on Parasitism of native Grasshoppers. Report of the Entomology Society of Ontario 70: 57-62.
Smith, R.W., and T.U. Finlayson. 1950. Larvae of dipterous parasites of nymphal and adult grasshoppers. Canadian Journal of Research 28: 81-117.
Streett, D.A., and M.R. McGuire. 1990. Pathogenic diseases of grasshoppers. Pp. 483-516 In: R.F. Chapman and A. Joern, A. (eds). Biology of grasshoppers. John Wiley & Sons.
Tourism Îles de la Madeleine. 2016a. Accessed September 8, 2016.
Tourism Îles de la Madeleine. 2016b. Un bilan 2012 positif pour Tourisme Îles de la madeleine . Accessed September 8, 2016.
Vickery, V. R., and D.K.M. Kevan. 1985. The grasshopper, crickets, and related insects of Canada and adjacent regions. Biosystematics Research Institute, Ottawa, Ontario. Publication Number 1777. 918 pp.
Vickery, V.R. 1989. The biogeography of Canadian Grylloptera and Orthoptera. Canadian Entomologist 121: 389-424.
Vickery, V.R., and D.K.E. Kevan. 1978. A new species of Melanoplus (Orthoptera: Acrididae: Melanoplinae) from the Magdalen Islands, Québec. Annals of the Entomological Society of Québec 22: 188-192.
Wood, J. 1973. Arthropoda which have been found in starling (Sturnus vulgaris) faeces. New Zealand Entomologist 5: 159-162.
York, G.T., and H.W. Prescott. 1952. Nemestrinid parasites of grasshoppers. Journal of Economic Entomology 45: 5-10.
Biographical summary of report writers
Cory S. Sheffield has been studying bees and pollination since 1993, as part of undergraduate honours studies at Acadia University, Wolfville, Nova Scotia. He continued graduate studies (MSc) of insect-plant interactions at Acadia, and at Agriculture and Agri-Food Canada (AAFC), Kentville, Nova Scotia from 1994 - 2006. Cory did graduate studies (PhD) at the University of Guelph, Ontario, while continuing to work at the AAFC. These studies focused on the bee fauna of Nova Scotia, including their diversity and contributions to crop pollination. During this time, Cory and several co-authors published on the re-discovery of Epeoloides pilosulus in Nova Scotia, which was thought extinct. Cory then worked on post-doctoral studies at York University, ON in bee taxonomy and DNA barcoding, followed by a research associate position in bee taxonomy with the Canadian Pollination Initiative (CANPOLIN). He is now research scientist and curator of invertebrate zoology at the Royal Saskatchewan Museum in Regina, SK. His research continues to focus on bees: he has published on the taxonomy of Canadian/North American bees, the utility of DNA barcoding for bees, bee physiology, pollination contributions and diversity of the Canadian bee fauna.
Jennifer Heron has been working on invertebrate conservation topics beginning with her MSc in 1998. She is currently the provincial invertebrate specialist with the B.C. Ministry of Environment. She directs and manages the provincial approach to invertebrate conservation, including the development and implementation of provincial legislation, policy, procedures, and standards for the conservation, and recovery of invertebrate species at risk, their habitats and ecosystems, and to keep these species from becoming at risk. She works with other invertebrate specialists to develop recovery-planning approaches and assign conservation status ranks to invertebrate groups. She works with local conservation and stewardship groups to achieve common public outreach goals.
Collections examined
Canadian National Collection of Insects, Arachnids and Nematodes, Ottawa, Ontario.
Lyman Museum, McGill University, Montréal, Québec.
Page details
- Date modified: